
12. tbl. 101. árg. 2015
Fræðigrein
Straumhvörf í rannsóknum á fjölhæfum stofnfrumum og notagildi þeirra í læknavísindum
Breakthrough in research on pluripotent stem cells and their application in medicine
Ágrip
Stofnfrumur úr fósturvísum eru einangraðar úr fósturvísum eins og nafnið bendir til. Þetta eru fjölhæfar frumur sem geta annaðhvort endurnýjast og haldist ósérhæfðar eða sérhæfst í hvaða frumugerð sem er í líkamanum. Árið 1998 tókst að einangra stofnfrumur úr fósturvísum manna og breytti það sýn manna á nýja möguleika í vefjalæknisfræði. Aðeins 8 árum síðar tókst vísindafólki að mynda svokallaðar iPS-frumur, fjölhæfar stofnfrumur sem útbúnar voru með því að endurforrita líkamsfrumur. Þetta hefur gjörbylt hugmyndum um óafturkræfi frumuþroska. Í kjölfarið hefur mikið verk verið unnið til þess að kryfja til mergjar sameindalíffræði fjölhæfra stofnfrumna. Unnt er að mynda iPS-frumur úr líkamsfrumum sjúklinga og hafa þær því sama genamengi. Þessar frumur eru því einstaklega nytsamar á ýmsum sviðum læknisfræðinnar og má meðal annars nýta þær til að skilja sjúkdómsframvindu, framkvæma lyfjaprófanir og vefjaígræðslur.
Stofnfrumur úr fósturvísum manna – hES-frumur
Stofnfrumur hafa tvenns konar eiginleika: við frumuskiptingu geta þær annaðhvort endurnýjað sig eða sérhæfst í aðra frumugerð. Stofnfrumur úr fósturvísum (ES-frumur, embryonic stem cells) eru einangraðar úr fósturvísum. Þær eru fengnar úr glasafrjóvgunum með upplýstu samþykki aðstandenda og þeim hefði annars átt að farga.1Þær teljast fjölhæfar (pluripotent) því þær geta sérhæfst í hvaða frumugerð líkamans sem er, ólíkt vefjasértækum stofnfrumum (tissue specific stem cells) sem geta aðeins sérhæfst í tiltekna frumugerð í vefnum sem þær finnast í, og eru því kallaðar marghæfar (multipotent).2 Í gegnum tíðina hafa stofnfrumur úr fósturvísum verið ranglega nefndar fósturstofnfrumur. Í fóstri eru engar fjölhæfar stofnfrumur heldur aðeins vefjasértækar stofnfrumur. Margir renna hýru auga til ES-frumna sem úrræðis til lækninga á ýmsum sjúkdómum, þar sem hægt verður að sérhæfa þær í ákveðnar frumugerðir sem sjúklingar þurfa á að halda, og væri þá hægt að koma þeim frumum fyrir í sjúklingnum. Sá böggull fylgir þó skammrifi að vefjaflokka-prótein eru tjáð í ES-frumunum og gæti það orsakað höfnun frumnanna við ígræðslu.3
Fósturþroskun mannsins
Okfruma myndast við samruna eggs og sáðfrumu. Hún skiptir sér á fyrstu stigum fósturþroska og verður að hneppifóstri (morula) á 16. frumustigi. Kímblaðran (blastocyst) myndast svo á 4-5 degi og innheldur hún innri frumumassa (inner cell mass) umlukinn ytra lagi af næringarhýði (trophectoderm) sem síðar þroskast í utanfósturvefi, svo sem fylgjuna. Innri frumumassinn myndar kímþekju (epiblast) sem við myndun holfósturs (gastrulation) á 14. degi þroskast í fósturlögin þrjú, innlag, miðlag og útlag. Eitt af aðalsmerkjum holfóstursmyndunar er myndun frumrákarinnar og sérhæfing hinna þriggja fósturlaga.1Fjölgun og frumuskrið kímþekjufruma fer fram í ferli sem kallast bandvefsumbreyting þekjuvefjar (Epithelial-to-mesenchymal transition, EMT) en þá minnkar viðloðun milli frumna og tenginga þeirra við grunnhimnu og frumuskrið á sér stað. ES-frumur manna (hES-frumur) ganga einnig í gegnum EMT þegar þær sérhæfast í rækt. Við upphaf sérhæfingar eykst tjáning á EMT-stjórnpróteinum, svo sem SNAIL, SLUG og TWIST, en tjáning á viðloðunarpróteininu E-CADHERIN þverr.4-6
Ef ytra lagið er varlega skilið frá innri frumumassa á kímblöðrustigi og frumum innri frumumassans sáð á ræktunarskálar við rétt skilyrði þá vaxa upp frumur í eyjum (kóloníum) sem nefnast stofnfrumur úr fósturvísum (mynd 1). Langt er síðan því var spáð að hægt yrði að einangra hES-frumur. Þroskunarfræðingar höfðu rannsakað furðuæxli (teratocarcinoma) sem finnast í eista og legi og innihalda misleita vefjablöndu, svo sem hár, vöðva, bein og tennur. Þroskunarfræðingar fundu út að ef þeir fjarlægðu fósturvísa úr legi og sprautuðu þeim undir skinn á músum fóru að vaxa furðuæxli. Þeir uppgötvuðu líka að þessi æxli innihéldu ósérhæfðar stofnfrumur sem gátu sérhæfst í ólíkar frumugerðir háð umhverfinu og kölluðu þær EC-frumur(embryonal carcinoma). Það má því segja að stofnfrumurannsóknir hafa notið góðs af þeirri gríðarlegu þekkingu sem áunnist hafði með EC-frumunum. Árið 1981 tókst að einangra stofnfrumur úr fósturvísum músa7,8 en það var ekki fyrr en 19959 sem tókst að einangra slíkar frumur úr prímötum og þremur árum síðar úr mönnum.10 Af augljósum ástæðum er ekki hægt að sannreyna svipgerð hES-frumna með því að búa til blendingsfóstur (chimeras) eins og mögulegt er í músum. Kenniprótein (markers) sem þróuð voru til að rannsaka EC-frumur reyndust því vel til að athuga ósérhæfðu hES-frumurnar. Skilgreining á hES-frumum byggist á þrennu: hES-frumur eru ódauðlegar því þær hafa telomerasa-virkni, hES-frumur tjá sértæk stjórnprótein og sértæk kenniprótein á yfirborði sínu, og hES-frumur mynda afleiður allra kímlaganna in vitro og furðuæxli in vivo.
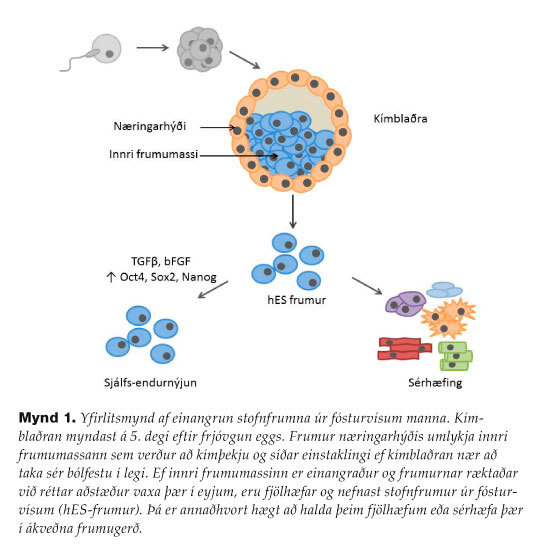{kind=link}
Aðferðir við stofnfrumurannsóknir
hES-frumur viðhalda fjölhæfi sínu þegar þær eru ræktaðar á geisluðum eða mitomycin-C meðhöndluðum MEF-hjálparfrumum (mouse embryonic fibroblasts). MEF-frumurnar geta því ekki skipt sér en seyta þó lífsnauðsynlegum þáttum og eru fótfesta fyrir stofnfrumurnar sem vaxa upp í eyjum. Á seinustu árum hefur komið í ljós að vaxtarþættirnir sem MEF-frumurnar seyta og halda hES-frumunum ósérhæfðum eru einkum bFGF (basic Fibroblast Growth Factor) og TGF-beta (Transforming Growth Factor beta).11,12 Þeir stuðla að tjáningu á stjórnpróteininu NANOG13 en heiti þessa próteins er myndað af orðunum Tír na nÓg sem þýðir land hinna ódauðlegu í keltneskum goðsögnum.14 NANOG ýtir undir tjáningu á OCT4 og SOX2 próteinunum en þessir þrír stjórnþættir eru aðalstjórnþættir hES-frumna og mynda eins konar hringrás hvað varðar stjórnun þeirra sín á milli (mynd 1).15,16 hES-frumum er vanalega umsáð með trypsíni eða svokallaðri cut-and paste-umsáningu og þær fluttar yfir á ferskt MEF-undirlag í ræktunarskálum. Sérhæfingu hES-frumna er einkum komið af stað með myndun frumuklasa (aggregates) sem í kjölfarið mynda frumukúlur (embryoid bodies) í fjarveru bFGF og TGF-beta. Þessar frumukúlur eru misleit blanda af alls kyns frumugerðum. Þetta kallast tilviljanakennd sérhæfing (spontaneous differentiation)17 en þó má beina sérhæfingu hES-frumna í tilteknar áttir með vaxtarþáttum eða hindrum (directed differentiation). Sem dæmi má nefna að hES-frumur sem eru örvaðar með FGF8, Sonic Hedgehog (SHH) og WNT-boðleiðinni en boðflutningur um TGF-beta og BMP (Bone Morphogenetic Protein) um leið hindraður, sérhæfast í dópamínmyndandi taugafrumur. Þetta hefur verið gert og frumum sem til verða sprautað í heila ungra músa með Parkinsonsjúkdóminn með árangri sem lofar góðu.18-20Örvun með bFGF og BMP4 (undirflokkur TGFbeta-fjölskyldunnar) leiðir til sérhæfingar í miðlagsfrumum sem svo er hægt að sérhæfa frekar í hjarta- og æðafrumur eða blóðmyndandi stofnfrumur.5,21 Ýmis dýralíkön eru notuð til að athuga öryggi, heimtur og starfhæfi þessara sérhæfðu frumna. Mýs, rottur og svín hafa til dæmis verið notuð til þess að græða hjartavöðvafrumur sem þannig voru búnar til í óstarfhæft hjarta eftir að hjartadrep var framkallað með því að binda fyrir kransæðarnar.22-24
Stuttu eftir að tókst að einangra stofnfrumur úr fósturvísum manna komu í ljós ýmis vandamál sem upp gætu komið í stofnfrumulækningum. Tryggja þarf að hES-frumurnar séu algerlega sérhæfðar í viðkomandi frumugerð, því ef hES-frumunum sjálfum væri sprautað í sjúkling gætu þær myndað furðuæxli. Í upphafi krafðist ræktun hES-frumna einnig MEF-hjálparfrumna auk kálfasermis og var þá hætta á veirumengun og jafnvel kúariðusmiti. Þessum ótta hefur verið eytt með því að sleppa MEF-hjálparfrumum og nota sermisfrítt æti. Ennfremur tjá hES-frumur vefjaflokka-prótein (HLA) á yfirborði sínu eftir sérhæfingu og leiðir það til mögulegrar höfnunar sjúklings á þessum frumum við ígræðslu.3 Ein lausn á þessu var að nota klónun, það er fjarlægja kjarna úr eggfrumu og setja kjarna úr líkamsfrumu sjúklings í eggfrumuna í staðinn. Þessi eggfruma yrði látin þroskast í kímblöðrustig, þar sem innri frumumassi yrði nýttur til sérhæfingar í tiltekna frumugerð sem væri nauðsynleg sjúklingnum. Hann myndi því ekki hafna frumum sem innhalda hans eigið genamengi. Slík klónun í mönnum í læknisfræðilegum tilgangi tókst nýlega.25
Rannsóknir á fósturvísum eru leyfðar á Íslandi, samanber lög um tæknifrjóvganir, nr. 55 frá 200826 og er því unnt að einangra stofnfrumur úr fósturvísum manna hér á landi samkvæmt núgildandi lögum. Þess ber þó að geta að slíkar rannsóknir eru undir ströngu eftirliti Vísindasiðanefndar og einangrun stofnfrumna úr mönnum hefur ekki verið framkvæmd hér á landi enn sem komið er, enda dýrt ferli.
Endurforritaðar frumur umturna kenningu um óafturkræfðar sérhæfðar frumur
Nóbelsverðlaunin í lífeðlis- og læknisfræði árið 2012 skiptust á milli tveggja vísindamanna, John B. Gurdon, Cambridge-háskóla, Bretlandi og Shinya Yamanaka, Kyoto-háskóla, Japan. Verðlaunin voru veitt fyrir að sýna að þroskaðar sérhæfðar frumur er hægt að endurforrita í fjölhæfar stofnfrumur. Niðurstöður þeirra breyttu þeirri almennu skoðun að sérhæfing líkamsfrumna (somatic cells) væri óafturkræf.27Gurdon klónaði fyrstur manna frosk þar sem hann flutti kjarna úr sérhæfðri líkamsfrumu úr froski í kjarnalaust egg, og sýndi fram á að erfðaupplýsingarnar úr líkamsfrumunni nægðu til að mynda halakörtu (1962). Yamanaka varð fyrstur til að umbreyta sérhæfðri líkamsfrumu beint í fjölhæfa stofnfrumu (2006).28
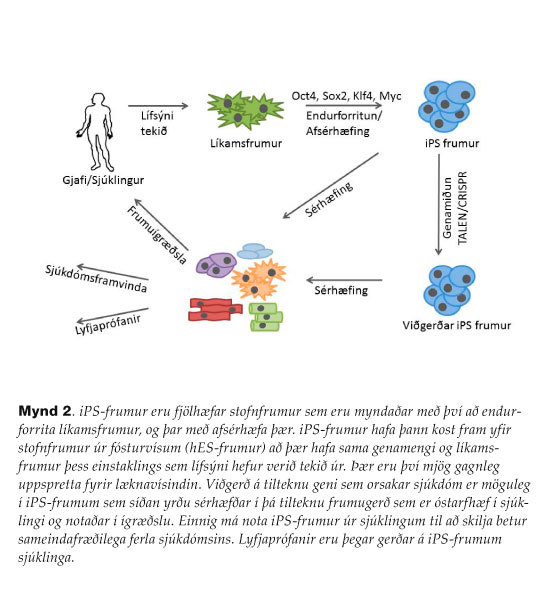
Í fósturþroskun eru fyrstu frumurnar í fósturvísi ósérhæfðar en verða svo sérhæfðari með tímanum. Í kringum 1950 höfðu þroskunarfræðingar klónað frosk með því að flytja kjarna úr fósturvísi í kjarnalaust froskaegg (Rana pipiens) og myndaðist þá fullmynduð halakarta (Somatic Cell Nuclear Transfer, SCNT).29Þegar þeir reyndu á hinn bóginn að endurtaka svipaða tilraun með kjarnaflutningi úr sérhæfðri frumu í kjarnalaust froskaegg, mistókst fósturþroskun í halakörtunni.30 Því var almennt talið að erfðaupplýsingar í sérhæfðum frumum gangist undir óafturkræfar breytingar sem gera það að verkum að frumurnar hafi takmarkaða sérhæfingarmöguleika. Þekkt myndlíking sem lýsti þessari viðteknu hugmynd vel var umframerfðalandslag (epigenetic landscape) Waddingtons sem sýndi kúlu uppi á fjallstoppi sem átti að líkjast ósérhæfðri stofnfrumu sem sérhæfðist meira og meira því lengra sem hún rúllaði niður fjallið og endaði að lokum í dalverpi sem sérhæfð fruma.31Niðurstöður John Gurdon breyttu þessari viðteknu skoðun manna. Hann notaði aðra froska-tegund (Xenopus laevis) í sínum rannsóknum. Honum tókst að flytja kjarna úr sérhæfðri líkamsfrumu frosksins (þekjufrumu meltingarvegarins) í kjarnalaust egg, og sýndi fram á að erfðaupplýsingarnar úr líkamsfrumunni nægðu til að mynda halakörtu, væru þær í réttu umhverfi. Þetta gerði hann árið 1962.32Með þessum niðurstöðum sýndi hann fram á að erfðaupplýsingar tapast ekki í sérhæfingarferlinu. Í dag vitum við að erfðaefnið er mistjáð í hinum mismunandi ósérhæfðu og sérhæfðu frumum. Gurdon breytti ekki aðeins grundvallarhugmyndum manna um óafturkræfni sérhæfðra frumna heldur innleiddi hann nýtt rannsóknarsvið og er stundum kallaður guðfaðir klónunar.33Spurningunni hvort hægt væri að umbreyta líkamsfrumu einni og sér beint í fjölhæfa stofnfrumu var þó enn ósvarað. Yamanaka hafði í nokkur ár verið að skoða hvaða stjórnprótein/umritunarþættir halda ES-frumum ósérhæfðum og fjölhæfum og uppgötvaði meðal annars að NANOG-stjórnpróteinið verður að vera tjáð svo að ES-frumur haldist fjölhæfar.34Þeir Takahashi og Yamanaka framkvæmdu síðan djarfa tilraun þar sem þeir komu mismunandi samsetningum af stjórngenum fyrir í bandvefsfrumum músa í frumurækt með hjálp retróveirusýkingar og notuðu valkvætt próf til að rekja þær frumur sem umbreyttust í fjölhæfar stofnfrumur. Þeim tókst að þrengja hringinn niður í aðeins fjögur stjórngen: OCT4, SOX2, KLF4 og MYC (sem nú eru nefndir Yamanaka-þættirnir) er gátu endurforritað líkamsfrumurnar í fjölhæfar stofnfrumur (mynd 2). Frumur þessar höfðu eiginleika sem voru fyllilega sambærilegir við eiginleika ES-frumna. Þessar frumur fengu nafnið iPS-frumur (induced Pluripotent Stem cells).28 Ári seinna tókst sama rannsóknarhópi að mynda iPS-frumur úr mönnum.35 Á fáum árum hefur orðið bylting í rannsóknum á iPS-frumum. Ýmsar ólíkar aðferðir hafa verið prófaðar við að mynda iPS-frumur, bæði hvað varðar innskeytingaraðferðir á stjórngenunum, svo og frumugerð líkamsfrumnanna. Einnig hafa færri stjórnþættir verið prófaðir og kemur í ljós að sleppa má MYC, enda hefur hann valdið æxlismyndunum í músum sem á kímblöðrustigi hafa verið sprautaðar með iPS-frumum. Þegar retróveirugenaferjum með fjölhæfigenunum er skeytt inn í frumurnar innlimast þær tilviljanakennt inn í genamengið og gætu því stuðlað að æxlismyndun. Þess vegna hefur verið reynt að komast hjá því að nota retróveirugenaferjur, til dæmis með veirugenaferjum sem innlimast ekki, með stöðugu RNA og með próteinum.36,37 Heimtur á fjölhæfum iPS-frumum eru lágar og því nauðsynlegt að nota valkvætt samrunagen til að velja úr þær frumur sem hafa endurforritast í iPS-frumur. Ferlið við að mynda iPS-frumur tekur um 2-3 vikur. En hvað gera Yamanaka-þættirnir til þess að ýta undir afsérhæfingu (de-differentiation)? Í stuttu máli virkja þeir gen sem taka þátt í fjölhæfi frumna en þagga aftur á móti niður í genum sem hafa hlutverki að gegna í frumusérhæfingu. Áður var greint frá að EMT á sér stað í fósturþroskun og við sérhæfingu hES-frumna. Yamanaka-þættirnir ýta þar af leiðandi undir endurforritun með öfugu ferli sem nefnist Mesenchymal-to-Epithelial transition (MET). Sýnt hefur verið að SOX2/OCT4 halda niðri EMT-stjórnpróteininu SNAIL, KLF4 ýtir undir tjáningu á E-CADHERIN og C-MYC hindrar TGF-β boðleiðina.38,39 Í framhaldinu eykst tjáning á NANOG, SOX2 og OCT4 genum frumunnar og þessi aðalstjórnprótein fjölhæfis valda virkjun á þeim genum sem nauðsynleg eru fjölhæfi, en virkja einnig svokölluð polycomb group prótein sem þagga niður tjáningu sérhæfingargena með litnisumbreytingu (chromatin remodelling).40
{kind=link}
Nytsemi fjölhæfra stofnfrumna
Frá læknisfræðilegu sjónarhorni gætu iPS-frumur orðið gagnlegar í lyfjaþróun og til skilnings á sjúkdómsferlum. Einnig væri hægt að hugsa sér að búa til sérhannaðar fjölhæfar stofnfrumur fyrir sjúklinga. Þessar frumur leysa nefnilega þann vanda að sjúklingur hafni ígræddum frumum enda hafa þær sama genamengi og sjúklingurinn sjálfur. Á seinustu árum hafa iPS-frumur verið búnar til úr afar mörgum sjúklingum og er aðgangur að upplýsingum þar að lútandi opinn.41
iPS-frumur sjúklinga hafa verið sérhæfðar í þá frumugerð sem sjúklingur hefur óstarfhæfa í líkama sínum og lyfjapróf gerð á þessum frumum. Raunverulegt dæmi um þessa nálgun er sameindafræðileg rannsókn á iPS-frumum úr barni sem þjáðist af hjartsláttartruflunum. Unnt var að greina stökkbreytingu í SCN5A-geninu sem olli longQT-heilkenni í barninu. Lyfjaprófanir á iPS-ættuðum hjartavöðvafrumum gerðu það að verkum að hægt var að meðhöndla barnið með réttum lyfjum.42 Lækning með iPS-frumum hefur sínar takmarkanir fyrir utan kostnaðarliðinn. Í fyrsta lagi þarf erfðaþátturinn að vega þungt í sjúkdómnum eins og til dæmis í arfgenga slímseigjusjúkdómnum cystic fibrosis, sem orsakast fyrst og fremst af einni stökkbreytingu í CFTR-geninu. Þann sjúkdóm má þá rannsaka í lungnaþekjufrumum. Á hinn bóginn væri erfitt að skoða sjúkdóma þar sem margir erfða- og umhverfisþættir spila saman. Í öðru lagi skipta viðmiðin öllu máli eins og í öðrum tilraunum. Þegar farið var að búa til iPS-frumur úr sjúklingum var notast við iPS-frumuviðmið úr heilbrigðum einstaklingum. Ólíkur erfðafræðilegur bakgrunnur í þessum frumum er mikið áhygguefni og gæti eins verið ástæða svipgerðarbreytinga í stað stökkbreytingarinnar sem verið er að rannsaka. Þetta er talið vera brotalöm á þessum rannsóknum.43 Í þriðja lagi er nauðsynlegt að iPS-frumur – bæði viðmið og þær sem hafa stökkbreytingu – séu búnar til á sama hátt, það er með nákvæmlega sömu aðferð við sömu ræktunaraðstæður. Til viðbótar má nefna mögulega skekkju vegna mismunandi umframerfða því það hefur sýnt sig að iPS-frumur hafa mismunandi metýleringarmynstur. Það getur haft áhrif á getu frumnanna til að sérhæfast í tiltekna frumugerð. Lausn á þessu viðmiðunarvandamáli er að nota fjölhæfar stofnfrumur sem hafa nákvæmlega sama erfðafræðilega bakgrunn utan stökkbreytingarinnar sem á að rannsaka, og mætti kalla þær samerfða (isogenic). Til þess að uppfylla slíkan gæðastaðal þarf annars vegar að mynda iPS-frumur úr sjúklingi sem hefur ákveðna stökkbreytingu og búa svo til viðmiðunar-iPS-frumur með því að leiðrétta sömu stökkbreytingu í sömu frumum. Að öðrum kosti eru notaðar hES-frumur sem viðmið en stökkbreyting sem veldur sjúkdómnum yrði þá mynduð í sömu frumum og svipgerð síðan athuguð. Þessi nálgun er nú viðtekin og sem dæmi um slíkar aðferðir má annars vegar nefna athugun á LRRK2-stökkbreytingu sem talin er valda Parkinsonsjúkdómi44 og rannsókn á stökkbreytingu í KCNH2 sem veldur longQT-heilkenni.45 Í Japan hefur löggjöf verið rýmkuð til þess að gera kleift að græða iPS-afleiddar litþekjufrumur í sjúkling sem þjáist af vota forminu af aldursbundinni hrörnun í augnbotnum. Ekki er búist við að í þessari fyrstu aðgerð sinnar tegundar verði strax mögulegt að lækna hrörnunina heldur frekar að koma í veg fyrir enn meiri hrörnun og athuga hliðarverkanir, svo sem ónæmissvörun eða æxlisvöxt samfara aðgerðinni.46,47
Þar sem ákveðin stökkbreyting í geni veldur sjúkdómnum þyrfti að leiðrétta stökkbreytinguna á iPS-frumustiginu með genamiðun (gene targeting) og sérhæfa frumurnar svo í kjölfarið í þá frumugerð sem sjúklingurinn þarf á að halda (mynd 2). Samstæð litningaendurröðun (homologous recombination) er mjög vel þekkt og áhrifarík aðferð til að slá út gen í músum („knockout-mýs“) en hún hefur reynst þung í vöfum við útslátt gena í hES-frumum og iPS-frumum.48 Á allra seinustu árum hafa miklar framfarir orðið í þessum geira. Transcription activator-like effector nucleases (TALENs) eru ensím sem kljúfa tvíþátta DNA á sértækum stöðum í genamenginu. Enn áhrifaríkari aðferð er CRISPR (Clustered regularly interspaced short palindromic repeats) tækni sem er ný aðferð sem leyfir breytingar á erfðaefni frumna og lífvera. Þessi tækni býður upp á afkastamikla aðferð í erfðatækni sem mætti nefna genaskurðlækningar. Nýlega eru nokkur dæmi þess að CRISPR-tæknin hafi verið notuð til að leiðrétta stökkbreyttar iPS-frumur úr sjúklingum, svo sem Duchenne-vöðvarýrnunarsjúkdóminn.49 Ýmsar framfarir hafa þegar orðið á þessari nýju tækni til þess að forðast „off-target“ áhrif og má gera ráð fyrir að aðferðin verði þróuð enn frekar á næstu árum.50-52
Stytt leið – Líkamsfrumum umbreytt beint í aðrar líkamsfrumur
Lengi hefur verið vitað að stjórnpróteinið MYOD1 stjórnar tjáningu gena sem hafa hlutverki að gegna í þroskun beinagrindavöðva. Ef þessu geni er skeytt í bandvefsfrumur þá virkjast vöðvaþroskunargen og frumurnar umbreytast í beinagrindavöðvafrumur.53 iPS-frumurnar ollu straumhvörfum í stofnfrumurannsóknum og í framhaldi af þessum mikla áfanga hafa vísindamenn prófað sig áfram með að umbreyta líkamsfrumum jafnvel beint í aðrar líkamsfrumur án milligöngu hinna fjölhæfu iPS-frumna (direct reprogramming). Ein af fyrstu tilraununum var gerð á rannsóknarstofu í sykursýki, þar sem briskirtilsfrumur voru endurforritaðar í insúlín-myndandi beta-frumur í músum in vivo. Níu genum sem eru þýðingarmikil í beta-frumuþroskun var skeytt inn í bris músar í mismunandi samsetningum. Í ljós kom að samsetning þriggja gena, sem eru umritunarþættirnirnir NGN3, PDX1 og MAFA, var nægileg og nauðsynleg til þess að stuðla að umbreytingu briskirtilsfrumna í beta-eyju-frumur (20% heimtur).54Þess má þó geta að báðar þessar frumugerðir eru afkomendur sama forverans og því um færri utanerfðaþætti að ræða heldur en við umbreytingu frumugerðar í aðra frumugerð sem jafnvel á uppruna í öðru fósturlagi. Í kjölfarið hefur mýmörgum frumum verið umbreytt. Vegna þess hversu algengt það er að hjarta- og æðasjúkdómar dragi fólk til dauða er mikilvægt að geta endurmyndað hjartavöðvafrumur. Fyrst tókst að umbreyta bandvefsfrumum í hjartavöðvafrumur með því að af-sérhæfa þær í átt að fjölhæfi en stöðva á ákveðnu millistigi (pluripotent intermediate) og sérhæfa þær þá í skilyrtu æti í hjartaforvera (mynd 3). Þetta hefur einnig verið gert með stjórnpróteinunum MESP1 og ETS2.55,56 Nú hefur tekist að umbreyta bandvefsfrumum beint í hjartavöðvafrumur in vitro með innskeytingu stjórnpróteinanna GATA4, MEF2C, TBX557 og HAND2.58Hjartadrep var framkallað í músum og stjórnprót-einum komið inn í bandvefsfrumur í skiptingu í örvefnum með hjálp retróveira en þær sýkja ekki hjartavöðvafrumur því þær eru ekki í skiptingu. Í tilraunum þessum var virkni hjartans bætt með hjálp umbreyttra bandvefsfrumna í starfhæfar hjartavöðvafrumur. Þótt hér hafi aðeins verið tekið dæmi um hjartavöðvafrumur hefur ýmsum líkamsfrumum (þó einkum bandvefsfrumum) verið umbreytt beint yfir í ýmsar frumugerðir.
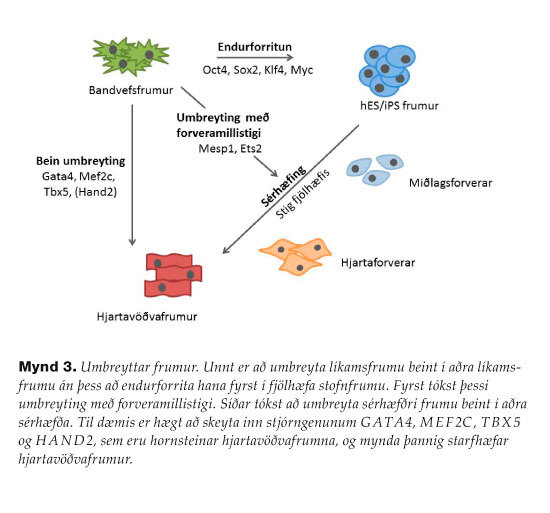{kind=link}
Samantekt
Í kjölfarið á fyrstu einangrun stofnfrumna úr fósturvísum manna (hES-frumum) árið 1998 urðu ótrúlegar framfarir í rannsóknum á þessum fjölhæfum stofnfrumum. Skilningur á sameindafræðilegum ferlum í fjölhæfum frumum jókst til muna og leiddi til þess að líkamsfrumur voru endurforritaðar í fjölhæfar stofnfrumur sem nefndar eru iPS-frumur og höfðu mjög svipaða eiginleika og hES-frumur. Þessi áfangi gjörbylti einnig hugmyndum fólks um óafturkræfi frumuþroska því niðurstöður tilraunarinnar sönnuðu að afsérhæfa mætti sérhæfðar frumur. Í framhaldinu hefur einnig tekist að umbreyta sérhæfðri frumu beint í aðra sérhæfða frumu af allt öðrum toga án fjölhæfa millistigsins. Miklar vonir eru bundnar við nytsemi fjölhæfra stofnfrumna í læknavísindum, það er í lyfjaprófunum og vefjaígræðslum. Nú þegar hafa verið framkvæmdar lyfjaprófanir á iPS-afleiddum frumum sjúklinga og þær gefið góða raun. Mikið kapp er lagt á að bæta aðferðir til þess að leiðrétta stökkbreytingar í iPS-frumum sjúklinga svo að hægt sé að græða iPS-afleiddar frumur aftur í sjúklinginn. Ýmsar hindranir eru á veginum, svo sem heimtur ígræddra frumna í skaddaðan vef sjúklings, auk þess sem slík einstaklingsbundin lækning yrði mjög kostnaðarsöm.
Þakkir
Ég vil þakka Eiríki Steingrímssyni og Ásdísi Kristjánsdóttur fyrir yfirlestur handrits og góðar ábendingar. Einnig vil ég þakka Ernu Magnúsdóttur fyrir gagnlegar samræður.
Heimildir
| 1. Pera MF, Trounson AO. Human embryonic stem cells: prospects for development. Development 2004; 131: 5515-25. http://dx.doi.org/10.1242/dev.01451 PMid:15509763 |
||||
| 2. Guðjónsson T, Steingrimsson E. Eiginleikar stofnfrumna: frumusérhæfing og ný meðferðarúrræði? Læknablaðið 2003; 89: 43-8. PMid:16819091 |
||||
| 3. Drukker M, Katz G, Urbach A, Schuldiner M, Markel G, Itskovitz-Eldor J, et al. Characterization of the expression of MHC proteins in human embryonic stem cells. Proc Natl Acad Sci U S A 2002; 99: 9864-9. http://dx.doi.org/10.1073/pnas.142298299 PMid:12114532 PMCid:PMC125045 |
||||
| 4. Eastham AM, Spencer H, Soncin F, Ritson S, Merry CL, Stern PL, et al. Epithelial-mesenchymal transition events during human embryonic stem cell differentiation. Cancer Res 2007; 67: 11254-62. http://dx.doi.org/10.1158/0008-5472.CAN-07-2253 PMid:18056451 |
||||
| 5. Richter A, Valdimarsdottir L, Hrafnkelsdottir HE, Runarsson JF, Omarsdottir AR, Ward-van Oostwaard D, et al. BMP4 promotes EMT and mesodermal commitment in human embryonic stem cells via SLUG and MSX2. Stem Cells 2014; 32: 636-48. http://dx.doi.org/10.1002/stem.1592 PMid:24549638 |
||||
| 6. Tan EJ, Olsson AK, Moustakas A. Reprogramming during epithelial to mesenchymal transition under the control of TGFbeta. Cell Adh Migr 2015; 9: 233-46. http://dx.doi.org/10.4161/19336918.2014.983794 PMid:25482613 PMCid:PMC4594534 |
||||
| 7. Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981; 292: 154-6. http://dx.doi.org/10.1038/292154a0 |
||||
| 8. Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci U S A 1981; 78: 7634-8. http://dx.doi.org/10.1073/pnas.78.12.7634 PMid:6950406 PMCid:PMC349323 |
||||
| 9. Thomson JA, Kalishman J, Golos TG, Durning M, Harris CP, Becker RA, et al. Isolation of a primate embryonic stem cell line. Proc Natl Acad Sci U S A 1995; 92: 7844-8. http://dx.doi.org/10.1073/pnas.92.17.7844 PMid:7544005 PMCid:PMC41242 |
||||
| 10. Thomson JA, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science 1998; 282: 1145-7. http://dx.doi.org/10.1126/science.282.5391.1145 PMid:9804556 |
||||
| 11. Xu RH, Peck RM, Li DS, Feng X, Ludwig T, Thomson JA. Basic FGF and suppression of BMP signaling sustain undifferentiated proliferation of human ES cells. Nat Methods 2005; 2: 185-90. http://dx.doi.org/10.1038/nmeth744 PMid:15782187 |
||||
| 12. Amit M, Itskovitz-Eldor J. Maintenance of human embryonic stem cells in animal serum- and feeder layer-free culture conditions. Methods Mol Biol 2006; 331: 105-13. http://dx.doi.org/10.1385/1-59745-046-4:105 |
||||
| 13. Xu RH, Sampsell-Barron TL, Gu F, Root S, Peck RM, Pan G, et al. NANOG is a direct target of TGFbeta/activin-mediated SMAD signaling in human ESCs. Cell Stem Cell 2008; 3: 196-206. http://dx.doi.org/10.1016/j.stem.2008.07.001 PMid:18682241 PMCid:PMC2758041 |
||||
| 14. Chambers I, Colby D, Robertson M, Nichols J, Lee S, Tweedie S, et al. Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 2003; 113: 643-55. http://dx.doi.org/10.1016/S0092-8674(03)00392-1 |
||||
| 15. Young RA. Control of the embryonic stem cell state. Cell 2011; 144: 940-54. http://dx.doi.org/10.1016/j.cell.2011.01.032 PMid:21414485 PMCid:PMC3099475 |
||||
| 16. Jaenisch R, Young R. Stem cells, the molecular circuitry of pluripotency and nuclear reprogramming. Cell 2008; 132: 567-82. http://dx.doi.org/10.1016/j.cell.2008.01.015 PMid:18295576 PMCid:PMC4142810 |
||||
| 17. Amit M, Itskovitz-Eldor J. Derivation and spontaneous differentiation of human embryonic stem cells. J Anat 2002; 200: 225-32. http://dx.doi.org/10.1046/j.1469-7580.2002.00032.x PMid:12033726 PMCid:PMC1570684 |
||||
| 18. Kriks S, Shim JW, Piao J, Ganat YM, Wakeman DR, Xie Z, et al. Dopamine neurons derived from human ES cells efficiently engraft in animal models of Parkinson's disease. Nature 2011; 480: 547-51. http://dx.doi.org/10.1038/nature10648 |
||||
| 19. Reubinoff BE, Itsykson P, Turetsky T, Pera MF, Reinhartz E, Itzik A, et al. Neural progenitors from human embryonic stem cells. Nat Biotechnol 2001; 19: 1134-40. http://dx.doi.org/10.1038/nbt1201-1134 PMid:11731782 |
||||
| 20. Kirkeby A, Grealish S, Wolf DA, Nelander J, Wood J, Lundblad M, et al. Generation of regionally specified neural progenitors and functional neurons from human embryonic stem cells under defined conditions. Cell Rep 2012; 1: 703-14. http://dx.doi.org/10.1016/j.celrep.2012.04.009 PMid:22813745 |
||||
| 21. Yang L, Soonpaa MH, Adler ED, Roepke TK, Kattman SJ, Kennedy M, et al. Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 2008; 453: 524-8. http://dx.doi.org/10.1038/nature06894 PMid:18432194 |
||||
| 22. Laflamme MA, Chen KY, Naumova AV, Muskheli V, Fugate JA, Dupras SK, et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat Biotechnol 2007; 25: 1015-24. http://dx.doi.org/10.1038/nbt1327 PMid:17721512 |
||||
| 23. Kehat I, Khimovich L, Caspi O, Gepstein A, Shofti R, Arbel G, et al. Electromechanical integration of cardiomyocytes derived from human embryonic stem cells. Nat Biotechnol 2004; 22: 1282-9. http://dx.doi.org/10.1038/nbt1014 PMid:15448703 |
||||
| 24. Van Laake LW, Van Hoof D, Mummery CL. Cardiomyocytes derived from stem cells. Ann Med 2005; 37: 499-512. http://dx.doi.org/10.1080/07853890500327843 PMid:16278163 |
||||
| 25. Tachibana M, Amato P, Sparman M, Gutierrez NM, Tippner-Hedges R, Ma H, et al. Human embryonic stem cells derived by somatic cell nuclear transfer. Cell 2013; 153: 1228-38. http://dx.doi.org/10.1016/j.cell.2013.05.006 PMid:23683578 PMCid:PMC3772789 |
||||
| 26. Lög um tæknifrjóvgun og notkun kynfrumna og fósturvísa manna til stofnfrumurannsókna. Lagasafn. Íslensk lög 1. október 2009. | ||||
| 27. Rossant J, Mummery C. NOBEL 2012 Physiology or medicine: Mature cells can be rejuvenated. Nature 2012; 492: 56. http://dx.doi.org/10.1038/492056a PMid:23222608 |
||||
| 28. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006; 126: 663-76. http://dx.doi.org/10.1016/j.cell.2006.07.024 PMid:16904174 |
||||
| 29. Briggs R, King TJ. Transplantation of Living Nuclei From Blastula Cells into Enucleated Frogs' Eggs. Proc Natl Acad Sci U S A 1952; 38: 455-63. http://dx.doi.org/10.1073/pnas.38.5.455 PMid:16589125 PMCid:PMC1063586 |
||||
| 30. King TJ, Briggs R. Changes in the Nuclei of Differentiating Gastrula Cells, as Demonstrated by Nuclear Transplantation. Proc Natl Acad Sci U S A 1955; 41: 321-5. http://dx.doi.org/10.1073/pnas.41.5.321 |
||||
| 31. Waddington CH. The strategy of the gene. A discussion of some aspects of theoretical biology. Allen and Unwin Ltd, London 1957. | ||||
| 32. Gurdon JB. The developmental capacity of nuclei taken from intestinal epithelium cells of feeding tadpoles. J Embryol Exp Morphol 1962; 10: 622-40. PMid:13951335 |
||||
| 33. Kolata G. Clone: The Road To Dolly, And The Path Ahead. William Morrow and Company Inc, New York 1998. | ||||
| 34. Mitsui K, Tokuzawa Y, Itoh H, Segawa K, Murakami M, Takahashi K, et al. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell 2003; 113: 631-42. http://dx.doi.org/10.1016/S0092-8674(03)00393-3 |
||||
| 35. Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007; 131: 861-72. http://dx.doi.org/10.1016/j.cell.2007.11.019 PMid:18035408 |
||||
| 36. Gonzalez F, Boue S, Izpisua Belmonte JC. Methods for making induced pluripotent stem cells: reprogramming a la carte. Nat Rev Genet 2011; 12: 231-42. http://dx.doi.org/10.1038/nrg2937 PMid:21339765 |
||||
| 37. Chun YS, Byun K, Lee B. Induced pluripotent stem cells and personalized medicine: current progress and future perspectives. Anat Cell Biol 2011; 44: 245-55. http://dx.doi.org/10.5115/acb.2011.44.4.245 PMid:22254153 PMCid:PMC3254878 |
||||
| 38. Li R, Liang J, Ni S, Zhou T, Qing X, Li H, et al. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell 2010; 7: 51-63. http://dx.doi.org/10.1016/j.stem.2010.04.014 PMid:20621050 |
||||
| 39. Samavarchi-Tehrani P, Golipour A, David L, Sung HK, Beyer TA, Datti A, et al. Functional genomics reveals a BMP-driven mesenchymal-to-epithelial transition in the initiation of somatic cell reprogramming. Cell Stem Cell 2010; 7: 64-77. http://dx.doi.org/10.1016/j.stem.2010.04.015 PMid:20621051 |
||||
| 40. Polo JM, Hochedlinger K. When fibroblasts MET iPSCs. Cell Stem Cell 2010; 7: 5-6. http://dx.doi.org/10.1016/j.stem.2010.05.018 PMid:20621040 |
||||
| 41. Santostefano KE, Hamazaki T, Biel NM, Jin S, Umezawa A, Terada N. A practical guide to induced pluripotent stem cell research using patient samples. Lab Invest 2015; 95: 4-13. http://dx.doi.org/10.1038/labinvest.2014.104 PMid:25089770 |
||||
| 42. Terrenoire C, Wang K, Tung KW, Chung WK, Pass RH, Lu JT, et al. Induced pluripotent stem cells used to reveal drug actions in a long QT syndrome family with complex genetics. J Gen Physiol 2013; 141: 61-72. http://dx.doi.org/10.1085/jgp.201210899 PMid:23277474 PMCid:PMC3536519 |
||||
| 43. Musunuru K. Personalized genomes and cardiovascular disease. Cold Spring Harb Perspect Med 2015; 5: a014068. http://dx.doi.org/10.1101/cshperspect.a014068 PMid:25256177 |
||||
| 44. Reinhardt P, Schmid B, Burbulla LF, Schondorf DC, Wagner L, Glatza M, et al. Genetic correction of a LRRK2 mutation in human iPSCs links parkinsonian neurodegeneration to ERK-dependent changes in gene expression. Cell Stem Cell 2013; 12: 354-67. http://dx.doi.org/10.1016/j.stem.2013.01.008 PMid:23472874 |
||||
| 45. Bellin M, Casini S, Davis RP, D'Aniello C, Haas J, Ward-van Oostwaard D, et al. Isogenic human pluripotent stem cell pairs reveal the role of a KCNH2 mutation in long-QT syndrome. Embo J 2013; 32: 3161-75. http://dx.doi.org/10.1038/emboj.2013.240 PMid:24213244 PMCid:PMC3981141 |
||||
| 46. Cyranoski D. Japanese woman is first recipient of next-generation stem cells 2014. 15.03.2015; nature.com/news/japanese-woman-is-first-recipient-of-next-generation-stem-cells-1.15915 - apríl 2015. | ||||
| 47. Pilot safety study of iPSC-based intervention for wet-type AMD 2013. 12.03.2015; riken-ibri.jp/AMD/english/research/index.html - apríl 2015. | ||||
| 48. Moretti A, Laugwitz KL, Dorn T, Sinnecker D, Mummery C. Pluripotent stem cell models of human heart disease. Cold Spring Harb Perspect Med 2013; 3: 49. http://dx.doi.org/10.1101/cshperspect.a014027 PMid:24186488 PMCid:PMC3808770 |
||||
| 49. Li HL, Fujimoto N, Sasakawa N, Shirai S, Ohkame T, Sakuma T, et al. Precise Correction of the Dystrophin Gene in Duchenne Muscular Dystrophy Patient Induced Pluripotent Stem Cells by TALEN and CRISPR-Cas9. Stem Cell Reports 2015; 4: 143-54. http://dx.doi.org/10.1016/j.stemcr.2014.10.013 PMid:25434822 PMCid:PMC4297888 |
||||
| 50. Fu Y, Sander JD, Reyon D, Cascio VM, Joung JK. Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nat Biotechnol 2014; 32: 279-84. http://dx.doi.org/10.1038/nbt.2808 PMid:24463574 PMCid:PMC3988262 |
||||
| 51. Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, Zhang F. Genome engineering using the CRISPR-Cas9 system. Nat Protoc 2013; 8: 2281-308. http://dx.doi.org/10.1038/nprot.2013.143 PMid:24157548 PMCid:PMC3969860 |
||||
| 52. Tsai SQ, Wyvekens N, Khayter C, Foden JA, Thapar V, Reyon D, et al. Dimeric CRISPR RNA-guided FokI nucleases for highly specific genome editing. Nat Biotechnol 2014; 32: 569-76. http://dx.doi.org/10.1038/nbt.2908 PMid:24770325 PMCid:PMC4090141 |
||||
| 53. Davis RL, Weintraub H, Lassar AB. Expression of a single transfected cDNA converts fibroblasts to myoblasts. Cell 1987; 51: 987-1000. http://dx.doi.org/10.1016/0092-8674(87)90585-X |
||||
| 54. Zhou Q, Brown J, Kanarek A, Rajagopal J, Melton DA. In vivo reprogramming of adult pancreatic exocrine cells to beta-cells. Nature 2008; 455: 627-32. http://dx.doi.org/10.1038/nature07314 PMid:18754011 |
||||
| 55. Islas JF, Liu Y, Weng KC, Robertson MJ, Zhang S, Prejusa A, et al. Transcription factors ETS2 and MESP1 transdifferentiate human dermal fibroblasts into cardiac progenitors. Proc Natl Acad Sci U S A 2012; 109: 13016-21. http://dx.doi.org/10.1073/pnas.1120299109 PMid:22826236 PMCid:PMC3420197 |
||||
| 56. Efe JA, Hilcove S, Kim J, Zhou H, Ouyang K, Wang G, et al. Conversion of mouse fibroblasts into cardiomyocytes using a direct reprogramming strategy. Nat Cell Biol 2011; 13: 215-22. http://dx.doi.org/10.1038/ncb2164 PMid:21278734 |
||||
| 57. Qian L, Huang Y, Spencer CI, Foley A, Vedantham V, Liu L, et al. In vivo reprogramming of murine cardiac fibroblasts into induced cardiomyocytes. Nature 2012; 485: 593-8. http://dx.doi.org/10.1038/nature11044 PMid:22522929 PMCid:PMC3369107 |
||||
| 58. Song K, Nam YJ, Luo X, Qi X, Tan W, Huang GN, et al. Heart repair by reprogramming non-myocytes with cardiac transcription factors. Nature 2012; 485: 599-604. http://dx.doi.org/10.1038/nature11139 PMid:22660318 PMCid:PMC3367390 |
||||