
10. tbl. 101. árg. 2015
Fræðigrein
Kímfrumur manna. Framfarir í frumurækt og vonir um meðferðarúrræði
Ágrip
Kímfrumur eru forverar egg- og sáðfruma í mönnum, sem bera erfðaupplýsingar á milli kynslóða. Vegna þess að kímfrumur sérhæfast snemma á fósturskeiði, rétt við bólfestu fósturvísis í legslímhúð, eru þær óaðgengilegar til rannsókna. Þekking okkar á tilurð þeirra hefur því til þessa verið afar takmörkuð og að mestu byggð á rannsóknum á dýramódelum eins og músum og kanínum. Í kjölfar rannsókna á stofnfrumum úr fósturvísum og eiginleikum þeirra hefur nú tekist að sérhæfa frumkímfrumur (primordial germ cells) manna á skilvirkan hátt í vefjarækt út frá stofnfrumum fósturvísa. Samhliða hefur tekist að sérhæfa frumkímfrumur úr iPS-frumum (induced pluripotent stem cells) manna sem eru myndaðar við afsérhæfingu líkamsfruma. Í þessari yfirlitsgrein verður farið yfir stöðu þekkingar okkar á frumkímfrumum manna og rannsókna á fjölhæfi stofnfruma úr fósturvísum manna og músa, ásamt því að ræða mögulega nýtingu frumuræktarkerfis fyrir frumfrjófrumur í rannsóknum og meðferð á ófrjósemi og öðrum kímfrumutengdum sjúkdómum.
Inngangur
Frumkímfrumur eru forverar kynfruma í fósturþroska og þroskast að lokum yfir í egg og sáðfrumur. Frumkímfrumur eru því sérhæfðar frumur sem bera með sér erfða- og umframerfðaupplýsingar (epigenetic information) frá foreldri til afkvæmis. Þær eru hinn ævarandi hlekkur á milli kynslóða sem er nauðsynlegur afkomu tegundanna.
Frumkímfrumur manna eru óaðgengilegar til rannsókna og því hefur vitneskja okkar á ákvörðun þeirra og þroska þróast mjög hægt síðan þeim var fyrst lýst fyrir rétt rúmum 100 árum.1 Meginþorri þekkingar okkar á mannakímfrumum hefur því verið leidd af rannsóknum á ákvörðun og þroska frumkímfruma í öðrum spendýrum. Erfðafræðitilraunir á músum síðustu tvo áratugi hafa afhjúpað erfðaþætti sem nauðsynlegir eru fyrir ákvörðun kímfruma og hafa flestar þær niðurstöður, en þó ekki allar, leitt af sér hliðstæða þekkingu á mannakímfrumum. Einnig hafa frumulíffræðitilraunir á fósturvísum músa og stofnfrumum úr fósturvísum bæði manna og músa verið mikilsverður bakgrunnur þekkingaröflunar á mannakímfrumum, tilurð þeirra og þroska.
Rannsóknir á kímfrumum og forverum þeirra eru forsenda framfara í æxlunarlæknisfræði (reproductive medicine). Ófrjósemi eða „undirfrjósemi“ (sub-fertility) hrjáir um 10% allra para á Vesturlöndum og eru meðferðarúrræði að mestu takmörkuð við glasafrjóvgun.2 Forsenda nýrra meðferða við ófrjósemi er að rannsaka til hlítar sérhæfingu og þroskun kímfruma og skapa þannig betri þekkingargrundvöll til uppbyggingar meðferðarúrræða.
Eitt helsta markmið núverandi rannsókna kímfrumum manna miðar að því að unnt verði á skilvirkan hátt að búa til fullþroska kímfrumur á rannsóknarstofu með sérhæfingu stofnfruma. Nýlega urðu straumhvörf í þessum rannsóknum þar sem tókst að sérhæfa frumkímfrumur manna í rækt úr stofnfrumum fósturvísa annars vegar og „induced pluripotent“ stofnfrumum (iPS) hins vegar.3-5 Að hafa stjórn á öllum skrefum í þroskaferi kímfruma í tilraunaglasi, frá ákvörðun þeirra til myndunar kynfruma af báðum kynjum, myndi efla rannsóknir á eðlilegri kynfrumumyndun. Einnig myndi ný þekking skapast á þeim ferlum sem fara úrskeiðis í þroskun kynfruma og valda til dæmis ófrjósemi eða æxlisvexti. Slíkt rækturnarkerfi yrði undirstaða framfara í hjálparaðferðum við frjóvgun (assisted reproductive technologies).
Nóbelsverðlaunin í læknisfræði árið 2012 voru veitt Sir John Gurdon og Shinya Yamanaka, en þeir eru frumkvöðlar á sviði dýraklónunar og hugmynda um frumusérhæfingu annars vegar og iPS-frumutækninnar hins vegar.6,7 iPS-frumutæknin hefur vakið vonir um einstaklingsbundin meðferðarúrræði við sjúkdómum sem til þessa hafa reynst ill- eða ólæknanlegir. Með þessari tækni má taka líkamsfrumur [feitletruð orð vísa til orðalista á bls. 464] einstaklinga, eins og til dæmis húðfrumur, og snúa sérhæfingu þeirra við þannig að þær öðlist að flestu eða öllu leyti sömu eiginleika og stofnfrumur fósturvísa. Þær öðlast því eiginleika fjölhæfis og geta þroskast yfir í allar frumutegundir mannslíkamans.
iPS-tæknin er ómetanleg þegar kemur að því að búa til líkön til rannsókna á ákveðnum sjúkdómum. Þannig hefur húðfrumum einstaklinga með þrístæðan litning 21 verið breytt í taugafrumur með því að snúa þeim fyrst yfir í iPS-frumur og þroska þær svo stig af stigi yfir í taugafrumur með það að markmiði að rannsaka frumulíffræði Alzheimer-sjúkdómsins.8 Einnig hafa á svipaðan hátt verið gerðar iPS-frumur einstaklinga með Parkinsons- og Huntingtons-veiki9, og svo mætti lengi telja.
Nýting á frumuræktarkerfi fyrir kímfrumur yrði einkum tvíþætt. Annars vegar yrði mögulegt að rannsaka sameinda- og frumulíffræðilegar forsendur kímfrumuþroska og hvernig þær færu úrskeiðis með því að nýta til þess iPS-frumur úr einstaklingum með erfðagalla sem valda ófrjósemi. Hins vegar yrði mögulega unnt að nýmynda kynfrumur til glasafrjóvgunar út frá líkamsfrumum einstaklinga með því að snúa þroska þeirra fyrst við með iPS-tækninni, og mynda svo fullþroskaðar kynfrumur. Þá mætti mögulega græða kím-stofnfrumur (germ-line stem cells) í einstaklinga, til dæmis eftir lyfjameðferðir sem valda tapi á kynfrumum (gonadotoxic treatments). Í dag er þetta enn fjarlægur möguleiki, sem þó þokast nær með nýjustu uppgötvunum.2
Nýlegar niðurstöður sem sýndu að hægt væri að sérhæfa markvisst kímfrumur manna í rækt hafa því vakið vonir um að unnt verði á næstu árum að fylgja þessum niðurstöðum eftir og ná fram fullum þroska frumanna í rækt, frá sérhæfingu og endurstillingu umframerfðamarka (epigenetic marks) til framköllunar meiósu. Forsendur þessara nýju uppgötvana á ákvörðun mannakímfruma byggja fyrst og fremst á þekkingu okkar á frumkímfrumumyndun músa og samanburði við eðli stofnfuma úr fósturvísum manna og músa.10 Í þessari yfirlitsgrein verður leitast við að varpa ljósi á þekkingu okkar á ákvörðun og sérhæfingu frumkímfruma manna með nýjustu uppgötvanir að leiðarljósi en mikilvægt er að styðjast við þekkingu okkar á frumkímfrumum annarra spendýra, einkum músa og kanína.
Sérhæfing og þroski frumkímfruma.
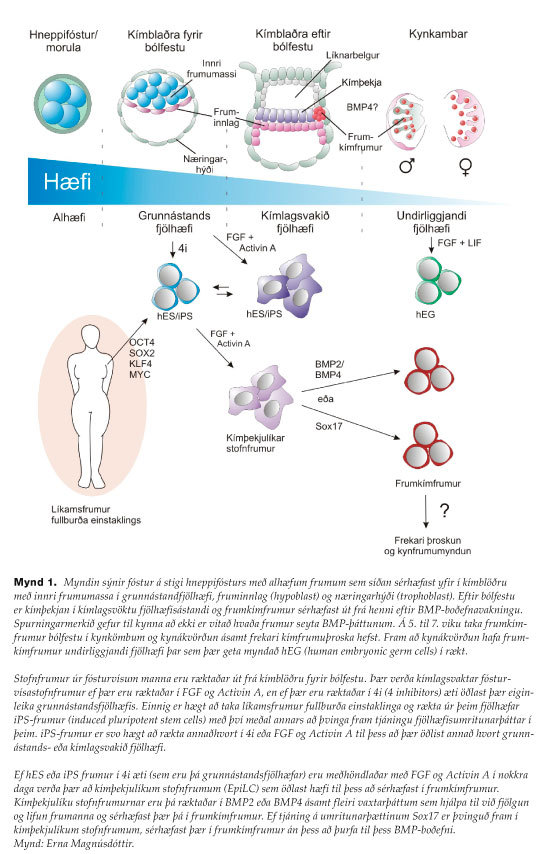
Kímblaðra fósturvísa tekur sér bólfestu í legþekju á annarri viku fósturþroska manna og myndun utanfóstursvefja hefst áður en holfóstursmyndun er hafin. Á þriðju viku fósturþroska myndast tvílaga diskur fósturvísisins þar sem kímþekjan (epiblast), sem samanstendur af fjölhæfum frumum sem munu gefa af sér öll kímlög fóstursins ásamt kímlínunni, liggur á milli fruminnlags (hypoblast) og líknarbelgs (mynd 1). Við upphaf fjórðu viku má finna fyrstu greinanlegu kímfrumur fósturvísisins í frumuþeli blómabelgs (yolk sac) við upphafstað frumrákarinnar (primitive streak), þar sem unnt er að aðgreina þær frá öðrum frumum með virkni Alkaline fosfatasa.11
{kind=link}
Frumkímfrumur eru því ákvarðaðar mjög snemma í fósturþroskanum, rétt fyrir holfóstursmyndun, samhliða fyrstu líkamsfrumusérhæfingu. Talið er að með því að setja til hliðar frumur til kynfrumumyndunar svo snemma í fósturþroska sé þeim forðað frá mögulegum erfðaefnisskemmdum sem eru fylgifiskur frumuskiptingar.
Í spendýrum eru frumkímfrumur ákvarðaðar með boðefnavakningu, þegar fjölhæfar frumur kímþekjunnar skynja boðefni frá nærlægum vefjum og öðlast þar með örlög kímlínunnar12 (mynd 1).
Frumkímfrumur spendýra ákvarðast því löngu áður en kynkirtlarnir verða til. Samhliða formþroskun fósturvísisins ferðast frumkímfrumurnar frá upprunastað sínum í kímþekju eftir afturgirni þar til þær ná kynkömbunum (genital ridges) sem eru forverar kynkirtlanna. Frumkímfrumurnar setjast að í kynkömbunum í lok 5. og byrjun 6. viku fósturþroskans. Samtímis þessu langa ferðalagi frumkímfrumanna fjölga þær sér og hefja endurstillingu umframerfðamarka með það að markmiði að afmá umframerfðaminni það sem greypt var með genagreypingu í erfðamengi kynfruma foreldranna.13-15 Einnig eru umframerfðamörk þau er mynduð voru í kímþelinu við undirbúning líkamsfrumusérhæfingar afmáð. Þessi endurstilling umframerfðamarka (epigenetic reprogramming) er mikilvæg til að ná grunnástandi umframerfðamarka (epigenetic ground state) áður en genagreyping er framkvæmd samkvæmt kyni fóstursins fyrir upphaf meiósu og kynfrumuþroskunar.
Þegar kynkömbunum er náð fjölga frumkímfrumurnar sér enn frekar. Á sama tíma eru þær umluktar strengjum líkamsfruma kynkambanna sem gegna lykilhlutverki við kynákvörðun. Á níundu viku fósturþroska er hægt að greina sérhæfingu eggstofnfruma (oogonia), meðal annars vegna hraða frumuskiptinga og aukins fjölda hvatbera. Frumuskipting eggstofnfruma fósturs varir fram á 11.-12. viku þegar þær hefja meiósuskiptingu sem lýkur ekki fyrr en við frjóvgun.1
Í karlkyns kynkömbum eru frumkímfrumur umluktar Sertoli-frumum og hafa nú stækkaða kjarna og greinanleg kjarnakorn. Eftir 9. viku eru sáðstrengir (seminiferous sex chords) greinanlegir og frumuskipting er mjög virk í karlkyns kynkömbum á fyrstu þremur mánuðum meðgöngu. Frumkímfrumur karlkyns fóstra á þessu stigi halda einnig áfram að tjá gen sem einkenna frumkímfrumur, en eggstofnfrumur hætta að tjá þau við upphaf kynákvörðunar. Á öðrum þriðjungi meðgöngu hættir fjölgun karlkyns frumkímfruma smám saman og tjáning frumkímfrumu- og fjölhæfisþátta stöðvast. Á 18. viku hafa nær allar kímfrumurnar hætt mítósuvirkni og kallast nú sáðstofnfrumur sem eru frumur í hvíldarfasa. Sáðstofnrumur eru hamlaðar frá því að hefja sáðfrumumyndun og meiósu fram að kynþroska.2
Forsendur framfara í ræktun kímfruma
Að baki þeim framförum í frumuræktun kímfruma manna sem nú hafa orðið liggja grundvallarspurningar sem tekist hefur að svara á undanförnum áratugum með erfða- og frumulíffræðirannsóknum á músum og stofnfrumum úr fósturvísum músa og manna.10
Í fyrsta lagi þurfti að svara því hvaða frumur væru forverar frumkímfruma og hefðu því hæfi til þess að sérhæfast í kímfrumur.16 Til þess að geta ræst sérhæfingu kímfruma í frumurækt var lykilatriði að hafa réttar forverafrumur við upphaf sérhæfingar. Í öðru lagi þurfti að finna hvaða boðefni ræstu sérhæfinguna og að skilgreina þá umritunarþætti sem miðla virkni boðefnanna og keyra áfram kímfrumuörlög.17-19
Þótt kímfrumur verði til á mjög afmörkuðum stað í kímþekju fósturvísis hafa allar kímþekjufrumurnar hæfi til þess að ákvarðast sem kímfrumur.20,21 Staðsetning kímfrumusérhæfingar afmarkast af staðsetningu boðefnaseytingar aðlægra vefja. Fjölhæfar frumur á mismunandi stöðum kímþekjunnar eru því jafngildar og hafa hæfileika til þess að mynda frumkímfrumur verði þær á vegi réttra boðefna.17,20 Þau boðefni sem gegna lykilhlutverki í að beina kímþekjufrumum í sérhæfingu frumkímfruma eru svokallaðir „bone morphogenetic protein“ (BMP) vaxtarþættir sem er seytt af aðlægum frumum utanfóstursvefja.19,22-24 Tilraunir í músum leiddu í ljós tap á frumkímfrumum ef þessi gen eða viðtaka þeirra vantar, sem og umritunarþættina SMAD1 og SMAD5, sem miðla skilaboðum BMP-vaxtarþáttanna og berast inn í kjarna til að virkja genatjáningu einkennandi fyrir frumkímfrumur. Ef kímþekja manna eða músafósturvísa er sett í rækt ásamt háum styrk BMP4 eða BMP2 sérhæfast frumur upprunnar úr allri kímþekjunni í frumkímfrumur. Þessar tilraunir lögðu grundvöllinn að því að unnt væri að rækta frumkímfrumur músa og síðar manna á skilvirkan hátt in vitro með því að þroska þær fyrst yfir í kímþekjufrumur út frá stofnfrumum úr fósturvísum og rækta þær síðan í návist BMP4.16,25,26
Rannsóknir á fjölhæfum stofnfrumum úr fósturvísum hafa haldist þétt í hendur við rannsóknir á frumkímfrumum spendýra, ekki síst vegna þess að frumkímfrumur tjá umritunarþættina OCT4, SOX2 og NANOG sem nauðsynlegir eru í innri frumumassa kímblöðrunnar og stofnfrumum úr fósturvísum. Þessir þættir viðhalda fjölhæfi frumanna jafnframt því sem þeir ræsa fjölhæfisprógram í líkamsfrumum og stuðla að myndun iPS-fruma.6 Einnig eru frumur kímþekjunnar, sem eru forverafrumur kímfruma, fjölhæfar og svipar mjög til stofnfruma úr fósturvísum. Þannig er tjáningu fjölhæfisgena haldið við í frumkímfrumum við sérhæfingu þeirra á meðan hún er bæld í sérhæfingu líkamsfruma sem á sér stað samhliða sérhæfingu kímfruma.27
Þrátt fyrir tjáningu fjölhæfisþátta eru frumkímfrumur sérhæfðar frumur sem aðeins gefa af sér kímfrumur og teljast því hafa undirliggjandi fjölhæfi. Þetta fjölhæfi er meðal annars afhjúpað þegar kímfrumur manna eru settar í vefjarækt ásamt „fibroblast growth factor“ (FGF) og „leukemia inhibitory factor“ (LIF). Við það verða frumurnar aftur fjölhæfar og er hægt að viðhalda þeim í rækt ásamt því að unnt er að sérhæfa þær í frumur fósturlaganna þriggja.28 Þessar frumur eru kallaðar „embryonic germ cells“ (EG) og svipar mjög til stofnfruma úr fósturvísum að eðliseiginleikum, meðal annars vegna þess að hægt er að setja þær inn í kímblöðru músa þar sem þær þroskast samhliða öðrum frumum innri frumumassa kímblöðrunnar. Ólíkt EG-frumum músa þá viðhalda EG-frumur manna genagreypingu og þroskast því í öll kímlögin í músakímblöðrum sem komið er fyrir í ónæmisbældum hýslum, og mynda ekki sjálfkrafa æxli.29 EG-frumur teljast ekki lengur til kímlínunnar heldur hafa afsérhæfst og misst tjáningu gena sem tjáð eru í kímfrumum samhliða því að þær verða fjölhæfar.
Jafnframt því að tjá fjölhæfisþætti, bæla frumkímfrumur tjáningu gena sem stuðla að sérhæfingu líkamsfruma.3,30 Umritunarþátturinn BLIMP1 er tjáður sértækt í frumkímfrumum við sérhæfingu þeirra og er nauðsynlegur fyrir myndun þeirra.31,32 Hann bælir tjáningu líkamsfrumugena svo sem Hox-gena og er því ábyrgur fyrir því að afmarka fjölhæfi kímfrumanna.33 Ásamt BLIMP1 eru tveir aðrir þættir, PRDM14 og AP2γ, ræstir á svipuðum tíma við sérhæfingu frumkímfruma músa. Þessir þrír þættir vinna saman við að keyra áfram sértæka genatjáningu einkennandi fyrir frumkímfrumur.34,35 Tap sérhvers þessara þátta veldur því að séhæfing frumkímfruma raskast.31,32,36,37
Þegar þessar rannsóknir úr músum voru heimfærðar upp á frumkímfrumur manna kom í ljós að þær tjá BLIMP1 og AP2γ en ekki PRDM14 við sérhæfingu.3-5 Einnig kom í ljós að mannafrumurnar tjáðu ekki fjölhæfisþáttinn SOX2 eins og músafrumurnar, heldur umritunarþáttinn SOX17 í staðinn,3,4,15 en sá þáttur er einnig mikilvægur í sérhæfingu innlags. Í mannakímfrumum er SOX17 því nauðsynlegt til þess að ræsa sérhæfingu frumkímfruma, því þegar SOX17-genið er slegið út í frumurækt myndast þær ekki.3 SOX17 ræsir því næst tjáningu á BLIMP1 ogAP2γ, en PRDM14 er aðeins dauft tjáð á þessu stigi.
Vegna þess að það tókst að sérhæfa frumkímfrumur manna í rækt, var mögulegt að staðfesta að margir þættir mikilvægir fyrir kímfrumusérhæfingu væru nauðsynlegir bæði í mönnum og músum. Einnig var afhjúpaður mikilvægur munur í verkun umritunarþátta á milli manna og músa og hann sannreyndur með útsláttartilraunum. Þessar tilraunir undirstrika því mikilvægi þess að koma á heildstæðu ræktarkerfi fyrir kímfrumur manna þannig að unnt verði að afhjúpa frekar mikilvæga stjórnþætti í þroskaferli þeirra.
Fjölhæfar stofnfrumur
Fyrstu fjölhæfu stofnfrumur úr fósturvísum músa (mES) voru einangraðar og ræktaðar árið 1981 af Gail Martin og Martin Evans og Matthew Kaufmann.58,59 Evans hlaut Nóbelsverðlaunin í lífeðlisfræði árið 2007 fyrir þessa uppgötvun sína en ræktun stofnfruma úr fósturvísum var meðal annars forsenda þess að unnt væri að erfðabreyta músum til notkunar í rannsóknum. Erfðabreytingar eru gerðar með því að breyta erfðaupplagi mES frumanna og koma þeim svo fyrir í kímblöðru fósturvísis. Erfðabreyttu frumurnar þroskast þá sem hluti af blendingsfóstri (chimera) og leggja til frumur í kímlínuna. Fram til ársins 1981 höfðu vísindamenn rannsakað fjölhæfi og frumusérhæfingu með því að nota svokallaðar „embryonal carcinoma“ eða EC-frumur, bæði úr mönnum og músum. EC-frumur eru taldar eiga uppruna sinn í kímfrumum, og eru æxlisfrumur sem geta sérhæfst í öll fósturlögin en leggja þó ekki til frumur til kímlínunnar í blendingsfóstrum vegna litningagalla sem hindra meiósu.
Árið 1998 fann svo James Thomson aðferð til þess að einangra og viðhalda fjölhæfum stofnfrumum úr fósturvísum manna (hES).60 Stuttu síðar voru svo embryonic germ cells (hEGs) einangraðar úr kímfrumum mannafóstra.29 Þær eru fjölhæfar og unnt að viðhalda þeim með Fibroblast growth factor (FGF) ásamt boðefninu LIF, sem einnig viðheldur fjölhæfi mES-fruma, og mEG-fruma, en ekki hES-fruma. Þegar hES-frumur eru ræktaðar í ákveðinni blöndu af boðefnahindrum sem kölluð er 4i (4 inhibitors), ná þær grunnástandsfjölhæfi og hafa hæfileika til þess að mynda kímfrumur, séu þær fyrst sérhæfðar yfir í kímþelslíkar frumur með kímlagsvakið fjölhæfi.41 Hugmyndafræðin á bak við 4i-ræktunaraðferðina við mannafrumurnar er svipuð og sú sem notuð er við 2i-kerfið til að halda músafrumunum í grunnástandsfjölhæfi, það er að koma í veg fyrir frumusérhæfingu og vakningu hennar, en hindrarnir sem notaðir eru í hvoru tilviki um sig eru þó mismunandi.
Tilbrigði við fjölhæfi: hæfnissvið kímfrumusérhæfingar
Forsendur þess að unnt var að sérhæfa frumkímfrumur manna í rækt var það innsæi sem fengist hefur á rannsóknum á eiginleikum fjölhæfra fruma í fósturvísum músa og mismunandi stofnfruma sem hafa verið ræktaðar úr þeim. Hér verður þessi innsýn í fjölhæfiseiginleika þessara mismunandi fruma útskýrð nánar.
Í fósturþroska er fjölhæfi skammvinnt ástand sem fylgir í kjölfarið á alhæfi okfrumunnar. Rannsóknir á mynstrum genatjáningar fjölhæfra fruma hafa leitt í ljós að mörg mismunandi stig fjölhæfis eru til og virðist fjölhæfið rúma samfellu af mismunandi genatjáningu.38,39
Okfruman hefur hæfi til þess að sérhæfast í allar frumugerðir lífverunnar, hvort sem um er að ræða líkamsfrumur, kímfrumur eða utanfósturvefi þá er mynda fylgjuna og aðra stoðvefi fóstursins og telst hún því vera alhæf. Við fyrstu frumusérhæfingu fósturþroskans verða til tvennskonar frumur, innri frumumassinn og næringarhýði. Innri frumumassinn er þyrping fjölhæfra fruma sem hafa hæfileika til þess að þroskast yfir í öll fósturlögin og eru hinar eiginlegu forverafrumur allra líkamsvefja auk kímlínunnar. Næringarhýðið leggur hins vegar engar frumur af mörkum til sjálfs fóstursins en umlykur innri frumumassann og styður hann og þroskast í utanfóstursvefi.
Við bólfestu fóstursins skiptist innri frumumassinn í kímþekju (primitive ectoderm/epiblast) og innlagskím (hypoblast) og í sömu andrá nær kímþekjan svokölluðu grunnástandsfjölhæfi sem einkennist af tjáningu á fjölhæfisumritunarþættinum NANOG ásamt OCT4, SOX2 og PRDM14, ásamt því að báðir X-litningar kvenkyns fóstra eru virkir. Við bólfestu fósturvísisins þróast fjölhæfi kímþekjunnar yfir í kímlagsvakið fjölhæfi sem er mjög skammvinnt ástand þar sem frumurnar bera enn þann eiginleika að geta þroskast í hvaða fósturlag sem er og byrja að þokast í átt að frekari þroska.40 Á þessu stigi öðlast frumur hæfileikann til sérhæfingar frumkímfruma, en á fyrri stigum fjölhæfis virðist þær skorta þann eiginleika.16,26
Stofnfrumur úr fósturvísum bæði manna og músa verða til þegar fósturvísir á kímblöðrustigi fyrir bólfestu fósturvísisins er settur í rækt. Munurinn á stofnfrumum úr fósturvísum músa og manna er þó sá að stofnfrumur músa líkjast frumum í grunnástandi fjölhæfis en mannafrumum svipar til kímlagsvakins fjölhæfisástands.39 Stofnfrumur fósturvísa músa svara boðefninu LIF til viðhalds fjölhæfisins og stofnfrumur fósturvísa manna svara þáttunum FGF og Activin.
Vegna þess að stofnfrumur úr fósturvísum músa tjá fjölhæfisþættina OCT4, SOX2 og NANOG, en vantar kímlínuþættina BLIMP1 og AP2γ, var talið að þvinguð tjáning þeirra síðarnefndu í stofnfrumum myndi ræsa kímlínusérhæfingu þeirra. Þá kom í ljós að frumurnar þoldu ekki tjáningu á BLIMP1 og dóu.34,35 Frumurnar svöruðu heldur ekki BMP-boðefnum með frumkímfrumusérhæfingu. Hæfnisástand forvera frumkímfruma var því enn ófundið. Lykillinn reyndist vera að ná fram eiginleikum kímlagsvakins fjölhæfis í stofnfrumum úr fósturvísum músa með tímabundinni rækt í FGF og Activin og líkja þannig eftir fósturþroskanum. Eftir það svöruðu músafrumurnar BMP4 annars vegar eða tjáningu á BLIMP1, AP2γ og PRDM14 hins vegar með frumkímfrumusérhæfingu.16,34,35 Með því að bæta boðefnunum LIF, Epidermal growth factor (EGF), Stem cell factor (SCF) og BMP8b fékkst skilvirk sérhæfing þar sem yfir 70% frumanna urðu að kímfrumum og hámörkun varð á afkomu frumanna17.
Þessar rannsóknir á músum juku bjartsýni á að unnt yrði að ná fram skilvirkri þroskun mannakímfruma í rækt með sömu boðefnum og umritunarþáttum. Það var þó ekki eins auðsótt og búist var við í fyrstu. Helsta ljónið í veginum var munurinn á sameindafræðilegu eðli fjölhæfis stofnfruma úr fósturvísum músa og manna.
Þótt stofnfrumum fósturvísa manna svipi mjög til forverafruma frumkímfruma (mynd 1) svara þær ekki BMP4 eða tjáningu umritunarþáttanna þriggja á sama hátt og músafrumurnar. Við ræktun í BMP4 sérhæfast þær þó í frumkímfrumur, en á mjög óskilvirkan hátt, þar sem innan við 1% frumanna sérhæfast. Þetta gefur til kynna að aðeins hluti frumanna sé inni á hæfnissviði sérhæfingar í frumkímfrumur.
Við sérhæfingu kímfruma músa var mikilvægt að rækta stofnfrumurnar í blöndu boðefnahindra sem kölluð er 2i „2 inhibitors“ til þess að tryggja einsleitni ræktanna og staðsetningu þeirra í grunnástandsfjölhæfi áður en kímlagsvakið fjölhæfi var framkallað.16,40 Þar sem grunnástandsfjölhæfi stofnfruma úr fósturvísum manna hafði fram að þessu aðeins verið þvingað með tjáningu umritunarþátta var ekki mögulegt að beita sömu aðferð fyrr en nokkru síðar þegar aðferð til framköllunar grunnástandsfjölhæfis í stofnfrumum úr fósturvísum manna var þróuð41–43.
Tímamót urðu í sérhæfingu frumkímfruma manna í rækt þegar unnt varð að framkalla kímlagsvakið fjölhæfi með ræktun grunnástandsfjölhæfra stofnfruma úr fósturvísum manna í FGF og Activin í tvo daga og gera þær þannig „kímþekjulíkar“ (Epiblast like cells –EpiLCs).3 Eftir þá meðferð svöruðu frumurnar á skilvirkan hátt boðefnunum BMP2 og BMP4 með því að mynda frumkímfrumur í rækt. Fyrsti björninn var unninn, því kerfisbundin og skilvirk sérhæfing frumkímfruma manna hafði tekist. Aðrir vísindamenn hafa síðan náð að þroska forvera miðlags úr hefðbundnum ræktum fósturvísafruma og tekist að ræsa frumkímfrumusérhæfingu eftir það4,5. Jafnframt tókst að meðhöndla iPS-frumur manna á sama hátt og kalla fram sérhæfingu þeirra í frumkímfrumur.
Fjölhæfi (pluripotency): Hæfileiki frumu til að þroskast í eða gefa af sér dótturfrumur sem mynda allar líkamsfrumur lífveru, sem og frumur kímlínunnar.
Alhæfi (totipotency): Hæfileiki frumu til þess að gefa af sér dótturfrumur sem mynda alla vefi lífveru, jafnt utanfóstursvefi eins og fylgju og aðra stoðvefi, sem og líkamsfrumur lífverunnar sjálfrar og kímlínu.
Líkamsfrumur (somatic cells): Allar frumur lífveru fyrir utan kímfrumur, sem mynda líkama lífverunnar. Líkamsfrumur og dótturfrumur þeirra geta ekki lagt frumur af mörkum til kímlínunnar og erfast því ekki til næstu kynslóðar.
Grunnástandsfjölhæfi (ground state pluripotency): Fjölhæfisástand sem kemur fyrir í frumum í innri frumumassa manna og músafóstra á sama tíma og fruminnlag (hypoblast) myndast. Í þessu ástandi er há tjáning fjölhæfisþátta, en lág eða engin tjáning umritunarþátta sem taka þátt í sérhæfingu í líkamsvefi.
Kímlagsvakið fjölhæfi (lineage primed pluripotency): Fjölhæfisástand fruma kímþekjunnar eftir bólfestu fóstursins, áður en þær sérhæfast í kímlögin þrjú eða frumkímfrumur. Frumur sem bera kímlagsvakið fjölhæfi hafa hafið tjáningu ákveðinna umritunarþátta sem stjórna vefjasérhæfingu en hafa þó ekki sérhæft sig eða skuldbundið sig til sérhæfingar.
Innri-frumumassi (inner cell mass, ICM): Fjölhæfur frumuklasi í kímblöðru sem þroskast yfir í kímþekju við bólfestu mannafósturs. Frumur innri frumumassans gefa af sér kímlögin þrjú auk kímfruma.
Kímþekja (epiblast): Fjölhæft þekjufrumulag fósturvísis eftir bólfestu sem gefur af sér kímlögin þrjú auk kímfruma.
Næringarhýði (trophectoderm): Ysta frumulag fósturvísis á stigi kímblöðru sem myndar utanfóstursvefi eins og fylgju.
Holfóstursmyndun (gastrulation): Kerfisbundin myndun þrílaga disks fósturvísis, sem samanstendur af innlagi, útlagi og miðlagi, úr frumum kímþekjunnar.
Formþroskun (morphogenesis): Þroskun fósturs og vefja í endanlega byggingu/lögun.
Kynkambar (genital ridge): Forverar kynkirtlanna sem þroskast úr fósturbandvef.
Genagreyping (genomic imprinting): Stjórnun á genatjáningu eftir samsætum þannig að samsætan sem erfist frá öðru foreldrinu er bæld en hin virk. Grundvöllur genagreypingar er DNA-metýlering sem er sett á stjórnraðir í kímfrumum fyrir meiósu. Fyrst er DNA-metýleringin sem erfðist frá foreldrum strokuð út og hún endurskrifuð eftir því af hvaða kyni fóstrið sjálft er.
Nýting frumkímfrumurækta til rannsókna á sjúkdómum
Brýn þörf er á að auka þekkingu okkar á þroska kynfruma. Ræktunarkerfi fyrir kímfrumur þar sem mögulegt er að hafa stjórn á öllum skrefum þroskaferilsins, frá sérhæfingu frumanna þar til þær fara í gegnum meiósu og kynfrumumyndun, myndi opna möguleika á því að svara rannsóknarspurningum sem snúa að mörgum eiginleikum kynkerfisins. Til dæmis myndi slíkt kerfi auðvelda til muna rannsóknir á ferlum kynákvörðunar, kynfrumuþroskunar og svo tilurð kímfrumukrabbameina og furðuæxla.44,45 Við rannsóknir á ófrjósemi yrði hægt að meðhöndla frumurnar í rækt, annað hvort með lyfjum eða gera erfðabreytingar á frumunum og skoða áhrif meðferðarinnar á þroskun frumanna í rækt. Þannig væri til dæmis mögulegt að skima fyrir efnum sem örva síðari stig kynfrumuþroska og líklegt væri að hefðu áhrif á frjósemi á fullorðinsárum. Einnig yrði mögulegt að gera erfðaskimanir fyrir þáttum sem hafa áhrif á kynfrumuþroskun.2
Fyrir meiósu eru genagreypingar frumkímfruma endurstilltar. Þannig er DNA-metýlering á genagreypingarsvæðum strokuð út og endurstillt eftir mynstri sem ákvarðast af því af hvoru kyninu fóstrið er. Verði villur í genagreypingu getur það haft alvarlegar afleiðingar fyrir fósturþroska, þar sem röng genagreyping getur valdið fósturgöllum og fósturláti. Röng genagreyping er til dæmis tengd sjúkdómum eins og Beckwith-Wiedermann-heilkenni, Rett-heilkenni og Wilm‘s æxli svo eitthvað sé nefnt.46 Nákvæm tímasetning endurstillingar genagreypingar er ekki þekkt í fóstrum manna, þó vitað sé að hún gerist síðar en í músum. Því er músamódelinu mjög ábótavant þegar kemur að rannsóknum á genagreypingu í mönnum og undirstrikar það þörfina fyrir ræktunarmódel kímfruma manna.47
Meðal annarra spurninga sem mögulegt væri að svara með ræktarmódeli fyrir kímfrumur væri tilurð svokallaðra carcinoma in situ (CIS) fruma, sem eru forverar eistnakímfrumuæxla (testicular germ cell tumours, TGCT) sem koma upp í ungum karlmönnum. CIS-frumur eru komnar af kímfrumum á fósturstigi sem hafa ekki þroskast eðlilega á stigi frumkímfruma eða gonocyta. Ekki er vitað hvaða ferlar liggja þar að baki, en CIS-frumur liggja í dvala í eistum í bernsku og þróun þeirra yfir í illkynja eistnafrumukrabbamein hefst ekki fyrr en eftir kynþroska. Þar sem mýs þróa ekki með sér CIS-frumur er ekki unnt að rannsaka tilurð þeirra í músamódelum. Við rannsóknir á tilurð CIS hefur til þessa verið notast við ígræðslu fruma úr mönnum í aðrar lífverur (xenograft), en ræktunarlíkan myndi auðvelda frumrannsóknir sem svo yrði hægt að staðfesta til dæmis með ígræðslu mannakímfruma í mýs eða rottur.48
Rannsóknir á iPS-frumum hafa nú þegar varpað ljósi á ástæður ófrjósemi í Turner-heilkenni. Þegar iPS-frumum úr einstaklingi með Turner-heilkenni var komið fyrir við eggjastokka músa þroskuðust þær yfir í kímfrumu-líkar frumur.49 Því er líklegt að X-litningurinn sé fremur nauðsynlegur fyrir viðhald kynfruma fram að fullorðinsárum en fyrir sjálfa kynfrumumyndunina. Getan til þess að sérhæfa frumkímfrumur úr iPS-frumunum mun nú líklega verða enn öflugra verkfæri til slíkra rannsókna.
Nýting rækta frumkímfruma til meðferðarúrræða
Helstu not á vefjaræktarkerfi kímfruma yrðu einkum til meðferðar á ófrjósemi. Í dag eru gjafaegg og -sæði notuð ásamt glasafrjóvgun þegar einstaklingar framleiða ekki eigin fullþroska kynfrumur. Stundum er þó mögulegt að frysta egg og sæði fyrir meðferð eins og lyfjameðferð við krabbameini. Þó er það ekki ávallt hægt, einkum ef um unga krabbameinssjúklinga er að ræða. Ef mögulegt væri að þroska að fullu iPS-frumur úr þessum einstaklingum yfir í kynfrumur, myndi þörfin á gjafakynfrumum minnka og gera fólki kleift að eignast börn sem væru erfðafræðilegir afkomendur þess. Þó verður að hafa í huga að iPS-frumum hefur líklega áskotnast meira af stökkbreytingum en venjulega verða í kímlínu manna. Því er ekki ljóst hversu örugg þessi aðferð muni reynast.
Til að meðhöndla ófrjósemi karla hafa verið bundnar vonir við að unnt verði að koma iPS-frumum eða iPS-afleiddum kímfrumum fyrir í eistum og framkalla þannig síðustu skrefin í sáðfrumuþroska í eistunum sjálfum þar sem rétta umhverfið er til staðar. Þannig mætti komast hjá tæknilegum hindrunum í vefjarækt. Þó hefur verið varað við prófunum á þessum aðferðum í mönnum, því jafnvel samgena (autologous) iPS-frumur sem komið hefur verið fyrir í eistnakirtlum músa hafa valdið ónæmishöfnun.50
Þótt tekist hafi að framkalla meiósu í karlkyns frumkímfrumum sem eru leiddar af iPS-frumum, hefur það þó aðeins verið gert með eldri aðferðum til að keyra áfram sérhæfingu frumkímfruma. Því voru hlutfallslega fáar frumur í ræktunum sem skiptu sér með meiósu.51 Ekki hefur tekist að þroska frumurnar frekar eftir meiósu úr forsáðfrumum (primary spermatocyte) yfir í sæðisfrumur.52 Það verður því áhugavert að fylgjast með rannsóknum næstu missera, þar sem skilvirk ræktun frumkímfruma mun auðvelda kerfisbundnar tilraunir á fullþroskun sáðfruma í rækt.
Nýlega tókst að þroska sáðstofnfrumur (spermatogonial stem cells) úr sjúklingum með launeista (cryptorchidism) í vefjarækt yfir í forsæðisfrumur (round spermatids) sem endurstilltu gagnagreypingu á réttan hátt og gátu frjóvgað eggfrumur músa eftir smásjárfrjóvgun (spermatid microinjection).53 Um 60% okfrumanna sem urðu til á þennan hátt fóru í gegnum fyrstu frumuskiptingu og töldust því hæfar til fósturþroska. Þessi framfaraskref benda til þess að ekki sé langt að bíða þar til unnt verði á skilvirkan hátt að framleiða sáðfrumur í tilraunaglasi með líkamsfrumur manna sem byrjunarefnivið.
Erfiðara hefur reynst að búa til eggfrumur í vefjarækt. Þótt tekist hafi að leiða eggfrumur músa út frá frumkímfrumum í rækt með því að koma þeim fyrir við eggjastokka til þroskunar áður en glasafrjóvgun var framkvæmd,54 eru engar rannsóknir sem lýsa álíka framförum í ræktun eggfruma manna. Í eggjastokkum eru ekki til staðar eggstofnfrumur eftir fæðingu og því ekki unnt að nota ræktun þeirra til rannsókna svipað og gert er við sáðstofnfrumur. Rannsóknir á tilurð eggfruma í rækt eru því tiltölulega skammt á veg komnar og mun líklega taka mun lengri tíma að ná því marki en tilurð sáðfruma.
Að lokum má nefna að tímamótaframfarir í erfðabreytingum urðu við uppgötvun og þróun erfðabreytingakerfis sem kallast CRISPR-Cas955 og hefur hlotið mikla athygli. Nú er unnt að erfðabreyta lífverum á mun skilvirkari hátt en áður, og tiltölulega auðvelt að framkvæma erfðabreytingar sem ekki skilja eftir nokkur önnur ummerki eða „ör“ í erfðaefninu55. Vísindasamfélagið hefur því kallað eftir umræðu um notkun þessarar tækni í tengslum við genalækningar (gene therapy).56 Hingað til hefur bann legið við því í mörgum löndum að erfðabreyta mannafrumum til notkunar við æxlun. Vegna tilkomu CRISPR-Cas9 tækninnar samhliða framförum í ræktun kímfruma er sá möguleiki ekki lengur fjarlægur að tæknilega mögulegt verði að gera við stökkbreytingar sem valda erfðasjúkdómum með erfðabreytingum á kímlínu manna á á öruggan hátt. Nú þegar hafa Bretar leyft notkun gjafa-hvatbera til genameðferðar á kímlínu.57 Verði öryggisskilyrði uppfyllt, virðist því ekki fjarri lagi að genalækningar á kímlínu manna verði leyfðar fyrr eða síðar. Þó er enn langt í land. Vísinda- og heilbrigðisstarfsmenn ásamt samfélaginu öllu verða að taka til þess afstöðu hvenær og hvort slíkar genalækningar séu ásættanlegur og æskilegur möguleiki innan þess siðferðisramma sem við setjum læknavísindunum.
Samantekt
Frumkímfrumur eru forverar kynfruma og mynda hinn ævarandi hlekk á milli kynslóða lífvera.
Torvelt hefur verið að rannsaka þroskun frumkímfruma manna vegna þess hversu óaðgengilegar þær eru á fósturþroska. Nú hefur tekist að sérhæfa frumkímfrumur manna með því að nota fjölhæfar fósturvísastofnfrumur og iPS-frumur sem upphafsfrumur í rækt. Því eru miklar vonir bundnar við að á næstu árum verði mögulegt að herma eftir öllum stigum kynfrumuþroska manna í vefjarækt, og þar með skapa grundvöll rannsókna á ófrjósemi og sjúkdómum tengdum þroskun kímfruma. Þannig eru líka bundnar vonir við að ræktun frumkímfruma geti valdið frekari framförum við þróun hjálparaðferða við frjóvgun og að unnt verði að nota ræktarkerfið til þess að varðveita frjósemi fólks eftir krabbameinsmeðferð.
Þakkir
Ég vil þakka Guðrúnu Valdimarsdóttur, Eiríki Steingrímssyni og Pétri Henry Petersen fyrir yfirlestur handrits og góðar athugasemdir og umræður.
Heimildir
| 1. De Felici M. Origin, Migration, and Proliferation of Human Primordial Germ Cells. In: Oogenesis. Springer London, London 2013: 19-37. http://dx.doi.org/10.1007/978-0-85729-826-3_2 |
|
| 2. Hayashi Y, Saitou M, Yamanaka S. Germline development from human pluripotent stem cells toward disease modeling of infertility. Fertil Steril 2012; 97: 1250-9. http://dx.doi.org/10.1016/j.fertnstert.2012.04.037 PMid:22656305 |
|
| 3. Irie N, Weinberger L, Tang WW, Kobayashi T, Viukov S, Manor YS, et al. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015; 160: 253-68. http://dx.doi.org/10.1016/j.cell.2014.12.013 PMid:25543152 PMCid:PMC4310934 |
|
| 4. Sasaki K, Yokobayashi S, Nakamura T, Okamoto I, Yabuta Y, Kurimoto K, et al. Robust In Vitro Induction of Human Germ Cell Fate from Pluripotent Stem Cells. Cell Stem Cell 2015; 17: 178-94. http://dx.doi.org/10.1016/j.stem.2015.06.014 PMid:26189426 |
|
| 5. Sugawa F, Araúzo-Bravo MJ, Yoon J, Kim KP, Aramaki S, Wu G, et al. Human primordial germ cell commitment in vitro associates with a unique PRDM 14 expression profile. EMBO J 2015; 34: 1009-24. http://dx.doi.org/10.15252/embj.201488049 PMid:25750208 |
|
| 6. Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006; 126: 663-76. http://dx.doi.org/10.1016/j.cell.2006.07.024 PMid:16904174 |
|
| 7. Gurdon JB. Adult frogs derived from the nuclei of single somatic cells. Dev Biol 1962; 4: 256-73. http://dx.doi.org/10.1016/0012-1606(62)90043-X |
|
| 8. Shi Y, Kirwan P, Smith J, MacLean G, Orkin SH, Livesey FJ. A Human Stem Cell Model of Early Alzheimer's Disease Pathology in Down Syndrome. Sci Transl Med 2012; 4: 124ra29-124ra29. | |
| 9. Park IH, Arora N, Huo H, Maherali N, Ahfeldt T, Shimamura A, et al. Disease-specific induced pluripotent stem cells. Cell 2008; 134: 877-86. http://dx.doi.org/10.1016/j.cell.2008.07.041 PMid:18691744 PMCid:PMC2633781 |
|
| 10. Magnúsdóttir E, Surani MA. How to make a primordial germ cell. Development 2014; 141: 245-52. http://dx.doi.org/10.1242/dev.098269 PMid:24381195 |
|
| 11. Chiquoine AD. The identification, origin, and migration of the primordial germ cells in the mouse embryo. Anat Rec 1954; 118: 135-46. http://dx.doi.org/10.1002/ar.1091180202 PMid:13138919 |
|
| 12. Extavour CG, Akam M. Mechanisms of germ cell specification across the metazoans: epigenesis and preformation. Development 2003; 130: 5869-84. http://dx.doi.org/10.1242/dev.00804 PMid:14597570 |
|
| 13. Guo F, Yan L, Guo H, Li L, Hu B, Zhao Y, et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015; 161: 1437-52. http://dx.doi.org/10.1016/j.cell.2015.05.015 PMid:26046443 |
|
| 14. Gkountela S, Zhang KX, Shafiq TA, Liao WW, Hargan-Calvopi-a J, Chen PY, et al. DNA Demethylation Dynamics in the Human Prenatal Germline. Cell 2015; 161: 1425-36. http://dx.doi.org/10.1016/j.cell.2015.05.012 PMid:26004067 |
|
| 15. Tang WW, Dietmann S, Irie N, Leitch HG, Floros VI, Bradshaw CR, et al. A Unique Gene Regulatory Network Resets the Human Germline Epigenome for Development. Cell 2015; 161: 1453-67. http://dx.doi.org/10.1016/j.cell.2015.04.053 PMid:26046444 PMCid:PMC4459712 |
|
| 16. Hayashi K, Ohta H, Kurimoto K, Aramaki S, Saitou M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 2011; 146: 519-32. http://dx.doi.org/10.1016/j.cell.2011.06.052 PMid:21820164 |
|
| 17. Ohinata Y, Ohta H, Shigeta M, Yamanaka K, Wakayama T, Saitou M. A signaling principle for the specification of the germ cell lineage in mice. Cell 2009; 137: 571-84. http://dx.doi.org/10.1016/j.cell.2009.03.014 PMid:19410550 |
|
| 18. Okamura D, Hayashi K, Matsui Y. Mouse epiblasts change responsiveness to BMP4 signal required for PGC formation through functions of extraembryonic ectoderm. Mol Reprod Dev 2005; 70: 20-9. http://dx.doi.org/10.1002/mrd.20136 PMid:15515057 |
|
| 19. Lawson KA, Dunn NR, Roelen BA, Zeinstra LM, Davis AM, Wright CV, et al. Bmp4 is required for the generation of primordial germ cells in the mouse embryo. Genes Dev 1999; 13: 424-36. http://dx.doi.org/10.1101/gad.13.4.424 PMid:10049358 PMCid:PMC316469 |
|
| 20. Tam PP, Zhou SX. The allocation of epiblast cells to ectodermal and germ-line lineages is influenced by the position of the cells in the gastrulating mouse embryo. Dev Biol 1996; 178: 124-32. http://dx.doi.org/10.1006/dbio.1996.0203 PMid:8812114 |
|
| 21. Lawson KA, Hage WJ. Clonal analysis of the origin of primordial germ cells in the mouse. Ciba Found Symp1994; 182: 68-91. PMid:7835158 |
|
| 22. Hayashi K, Kobayashi T, Umino T, Goitsuka R, Matsui Y, Kitamura D. SMAD1 signaling is critical for initial commitment of germ cell lineage from mouse epiblast. Mech Dev 2002; 118: 99-109. http://dx.doi.org/10.1016/S0925-4773(02)00237-X |
|
| 23. Ying Y, Zhao GQ. Cooperation of endoderm-derived BMP2 and extraembryonic ectoderm-derived BMP4 in primordial germ cell generation in the mouse. Dev Biol 2001; 232: 484-92. http://dx.doi.org/10.1006/dbio.2001.0173 PMid:11401407 |
|
| 24. Ying Y, Liu XM, Marble a, Lawson K a, Zhao GQ. Requirement of Bmp8b for the generation of primordial germ cells in the mouse. Mol Endocrinol 2000; 14: 1053-63. http://dx.doi.org/10.1210/mend.14.7.0479 PMid:10894154 |
|
| 25. Hayashi K, Ogushi S, Kurimoto K, Shimamoto S, Ohta H, Saitou M. Offspring from oocytes derived from in vitro primordial germ cell-like cells in mice. Science 2012; 338: 971-5. http://dx.doi.org/10.1126/science.1226889 PMid:23042295 |
|
| 26. Irie N, Tang WWC, Azim Surani M. Germ cell specification and pluripotency in mammals: a perspective from early embryogenesis. Reprod Med Biol 2014; 13: 203-15. http://dx.doi.org/10.1007/s12522-014-0184-2 PMid:25298745 PMCid:PMC4182624 |
|
| 27. Fuhrmann G, Chung AC, Jackson KJ, Hummelke G, Baniahmad A, Sutter J, et al. Mouse germline restriction of Oct4 expression by germ cell nuclear factor. Dev Cell 2001; 1: 377-87. http://dx.doi.org/10.1016/S1534-5807(01)00038-7 |
|
| 28. Shamblott MJ, Axelman J, Wang S, Bugg EM, Littlefield JW, Donovan PJ, et al. Derivation of pluripotent stem cells from cultured human primordial germ cells. Proc Natl Acad Sci U S A 1998; 95: 13726-31. http://dx.doi.org/10.1073/pnas.95.23.13726 PMid:9811868 PMCid:PMC24887 |
|
| 29. Turnpenny L. Derivation of Human Embryonic Germ Cells: An Alternative Source of Pluripotent Stem Cells. Stem Cells 2003; 21: 598-609. http://dx.doi.org/10.1634/stemcells.21-5-598 PMid:12968114 |
|
| 30. Saitou M, Barton SC, Surani MA. A molecular programme for the specification of germ cell fate in mice. Nature 2002; 418: 293-300. http://dx.doi.org/10.1038/nature00927 PMid:12124616 |
|
| 31. Robertson EJ, Charatsi I, Joyner CJ, Koonce CH, Morgan M, Islam A, et al. Blimp1 regulates development of the posterior forelimb, caudal pharyngeal arches, heart and sensory vibrissae in mice. Development 2007; 134: 4335-45. http://dx.doi.org/10.1242/dev.012047 PMid:18039967 |
|
| 32. Ohinata Y, Payer B, O'Carroll D, Ancelin K, Ono Y, Sano M, et al. Blimp1 is a critical determinant of the germ cell lineage in mice. Nature 2005; 436: 207-13. http://dx.doi.org/10.1038/nature03813 PMid:15937476 |
|
| 33. Kurimoto K, Yabuta Y, Ohinata Y, Shigeta M, Yamanaka K, Saitou M. Complex genome-wide transcription dynamics orchestrated by Blimp1 for the specification of the germ cell lineage in mice. Genes Dev 2008; 22: 1617-35. http://dx.doi.org/10.1101/gad.1649908 PMid:18559478 PMCid:PMC2428060 |
|
| 34. Magnúsdóttir E, Dietmann S, Murakami K, Günesdogan U, Tang F, Bao S, et al. A tripartite transcription factor network regulates primordial germ cell specification in mice. Nat Cell Biol 2013; 15: 905-15. http://dx.doi.org/10.1038/ncb2798 PMid:23851488 PMCid:PMC3796875 |
|
| 35. Nakaki F, Hayashi K, Ohta H, Kurimoto K, Yabuta Y, Saitou M. Induction of mouse germ-cell fate by transcription factors in vitro. Nature 2013; 501: 222-6. http://dx.doi.org/10.1038/nature12417 PMid:23913270 |
|
| 36. Yamaji M, Ueda J, Hayashi K, Ohta H, Yabuta Y, Kurimoto K, et al. PRDM14 ensures naive pluripotency through dual regulation of signaling and epigenetic pathways in mouse embryonic stem cells. Cell Stem Cell 2013; 12: 368-82. http://dx.doi.org/10.1016/j.stem.2012.12.012 PMid:23333148 |
|
| 37. Weber S, Eckert D, Nettersheim D, Gillis AJ, Schäfer S, Kuckenberg P, et al. Critical function of AP-2 gamma/TCFAP2C in mouse embryonic germ cell maintenance. Biol Reprod 2010; 82: 214-23. http://dx.doi.org/10.1095/biolreprod.109.078717 PMid:19776388 |
|
| 38. Yan L, Yang M, Guo H, Yang L, Wu J, Li R, et al. Single-cell RNA-Seq profiling of human preimplantation embryos and embryonic stem cells. Nat Struct Mol Biol 2013; 20: 1131-9. http://dx.doi.org/10.1038/nsmb.2660 PMid:23934149 |
|
| 39. Hackett JA, Surani MA. Regulatory Principles of Pluripotency: From the Ground State Up. Cell Stem Cell 2014; 15: 416-30. http://dx.doi.org/10.1016/j.stem.2014.09.015 PMid:25280218 |
|
| 40. Nichols J, Smith A. Naive and Primed Pluripotent States. Cell Stem Cell 2009; 4: 487-92. http://dx.doi.org/10.1016/j.stem.2009.05.015 PMid:19497275 |
|
| 41. Gafni O, Weinberger L, Mansour AA, Manor YS, Chomsky E, Ben-Yosef D, et al. Derivation of novel human ground state naive pluripotent stem cells. Nature 2013; 504: 282-6. http://dx.doi.org/10.1038/nature12745 PMid:24172903 |
|
| 42. Buecker C, Chen HH, Polo JM, Daheron L, Bu L, Barakat TS, et al. A murine ESC-like state facilitates transgenesis and homologous recombination in human pluripotent stem cells. Cell Stem Cell 2010; 6: 535-46. http://dx.doi.org/10.1016/j.stem.2010.05.003 PMid:20569691 PMCid:PMC3162213 |
|
| 43. Hanna J, Cheng AW, Saha K, Kim J, Lengner CJ, Soldner F, et al. Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs. Proc Natl Acad Sci U S A 2010; 107: 9222-7. http://dx.doi.org/10.1073/pnas.1004584107 PMid:20442331 PMCid:PMC2889088 |
|
| 44. MacLaughlin DT, Donahoe PK. Sex Determination and Differentiation. N Engl J Med 2004; 350: 367-78. http://dx.doi.org/10.1056/NEJMra022784 PMid:14736929 |
|
| 45. Stevens LC. Origin of testicular teratomas from primordial germ cells in mice. J Natl Cancer Inst 1967; 38: 549-52. PMid:6025005 |
|
| 46. Swales a. KE, Spears N. Genomic imprinting and reproduction. Reproduction 2005; 130: 389-99. http://dx.doi.org/10.1530/rep.1.00395 PMid:16183857 |
|
| 47. Hajkova P, Ancelin K, Waldmann T, Lacoste N, Lange UC, Cesari F, et al. Chromatin dynamics during epigenetic reprogramming in the mouse germ line. Nature 2008; 452: 877-81. http://dx.doi.org/10.1038/nature06714 PMid:18354397 PMCid:PMC3847605 |
|
| 48. Sharpe RM, Mitchell RT. The downside of "inappropriate messaging": New insight into the development of testicular germ cell tumours in young men? J Pathol 2013; 229: 497-501. http://dx.doi.org/10.1002/path.4167 |
|
| 49. Dominguez AA, Chiang HR, Sukhwani M, Orwig KE, Reijo Pera RA. Human germ cell formation in xenotransplants of induced pluripotent stem cells carrying X chromosome aneuploidies. Sci Rep 2014; 4: 6432. http://dx.doi.org/10.1038/srep06432 PMid:25242416 PMCid:PMC4170197 |
|
| 50. Zhao T, Zhang Z-N, Rong Z, Xu Y. Immunogenicity of induced pluripotent stem cells. Nature 2011; 474: 212-5. http://dx.doi.org/10.1038/nature10135 |
|
| 51. Vassena R, Eguizabal C, Heindryckx B, Sermon K, Simon C, van Pelt AM, et al. Stem cells in reproductive medicine: ready for the patient? Hum Reprod 2015; 30: 2014-21. http://dx.doi.org/10.1093/humrep/dev181 |
|
| 52. Eguizabal C, Montserrat N, Vassena R, Barragan M, Garreta E, Garcia-Quevedo L, et al. Complete meiosis from human induced pluripotent stem cells. Stem Cells 2011; 29: 1186-95. http://dx.doi.org/10.1002/stem.672 PMid:21681858 |
|
| 53. Yang S, Ping P, Ma M, Li P, Tian R, Yang H, et al. Generation of Haploid Spermatids with Fertilization and Development Capacity from Human Spermatogonial Stem Cells of Cryptorchid Patients. Stem Cell Reports 2014; 3: 663-75. http://dx.doi.org/10.1016/j.stemcr.2014.08.004 PMCid:PMC4223697 |
|
| 54. Hayashi K, Saitou M. Generation of eggs from mouse embryonic stem cells and induced pluripotent stem cells. Nat Protoc 2013; 8: 1513-24. http://dx.doi.org/10.1038/nprot.2013.090 PMid:23845963 |
|
| 55. Jinek M, East A, Cheng A, Lin S, Ma E, Doudna J. RNA-programmed genome editing in human cells. Elife 2013; 2013: 1-9. http://dx.doi.org/10.7554/elife.00471 |
|
| 56. Baltimore BD, Berg P, Botchan M, Carroll D, Charo RA, Church G, et al. Biotechnology. A prudent path forward for genomic engineering and germline gene modification. Science 2015; 348: 36–8. http://dx.doi.org/10.1126/science.aab1028 PMCid:PMC4394183 |
|
| 57. Callaway E. Reproductive medicine: The power of three. Nature 2014; 509: 414-7. http://dx.doi.org/10.1038/507414a http://dx.doi.org/10.1038/509414a |
|
| 58. Evans MJ, Kaufman MH. Establishment in culture of pluripotential cells from mouse embryos. Nature 1981; 292: 154-6. http://dx.doi.org/10.1038/292154a0 |
|
| 59. Martin GR. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc Natl Acad Sci 1981; 78: 7634-8. http://dx.doi.org/10.1073/pnas.78.12.7634 PMCid:PMC349323 |
|
| 60. Thomson J a, Itskovitz-Eldor J, Shapiro SS, Waknitz MA, Swiergiel JJ, Marshall VS, et al. Embryonic stem cell lines derived from human blastocysts. Science 1998; 282: 1145-7. http://dx.doi.org/10.1126/science.282.5391.1145 |
|