
Fræðigreinar
Ljósaðlögun keilna í kanínum og marsvínum
Ágrip
Tilgangur: Þegar sjónhimna er aðlöguð að ljósi er hægt að skrá sjónhimnurit (electroretinogram, ERG) keilna. Vitað er að við ljósaðlögun verða miklar breytingar í sjónhimnuriti keilna hjá mönnum, öpum og músum. Þessar tegundir hafa æðar í sjónhimnu. Í þessari rannsókn var athugað hvort sama eigi við um tegundir spendýra með lítið af æðum í sjónu (kanínur) eða engar (marsvín) æðar í sjónu og sem hafa tvær tegundir keilna en alls ólík sjónhimnurit í rökkri. Efniviður og aðferðir: Sjónhimnurit voru skráð frá svæfðum kanínum og marsvínum með skráningarskautum úr stálvír, sem staðsett voru á hornhimnu augans. Koparvír staðsettur í munni var notaður sem viðmiðunarskaut og nál undir húð sem jarðtenging. Skráning var mögnuð 1000-falt, með bandvídd 1-1000 Hz, og niðurstöður fluttar í tölvu með A/D breytikorti. Hornhimna var staðdeyfð með próparakaíni, sjáaldur víkkað með trópikamíð dropum. Sjónhimnurit var vakið með stuttum (10msek) ljósblikkum, og sjónhimna aðlöguð að stöðugu hvítu bakgrunnsljósi.
Niðurstöður: Í rökkri er b-bylgja meir en helmingi stærri en a-bylgja í sjónhimnuriti kanína, en b-bylgja aðeins eilítið stærri en a-bylgja hjá marsvínum. Í sjónhimnuriti keilna er hins vegar b-bylgja helmingi stærri en a-bylgja hjá báðum tegundum. Eftir að kveikt er á bakgrunnsljósi eftir aðlögun að rökkri hækkar spenna svara og nær hámarki eftir um 10 mínútur í báðum tegundum. Dvöl b-bylgju styttist hjá kanínum, en ekki hjá marsvínum. Í sjónhimnuriti keilna hjá kanínum eru sveifluspennur (oscillatory potentials), sem stækka að spennu við aðlögun að ljósi. Sveifluspennur eru í sjónhimnuriti marsvína, þegar það er skráð í rökkri, en engar í sjónhimnuriti keilna.
Ályktanir: Spendýr með æðalausa sjónhimnu og engar langbylgjukeilur sýna ljósaðlögun sjónhimnurits keilna. Í marsvínum eykst sjónhimnurit keilna að spennu án þess að dvöl breytist við ljósaðlögun. Þessar breytingar eru svipaðar í tíma í kanínum og marsvínum. Sveifluspennur aukast að spennu í kanínum en ekki marsvínum. Niðurstöður benda til að önnur ferli ráði ljósaðlögun sjónhimnurits keilna hjá marsvínum en kanínum.
English Summary |
| Möller A, Eysteinsson Þ Light adaptation of cones in rabbits and guinea pigs Læknablaðið 2001; 87: 221-6 Objective: During light adaptation of the retina, cone electroretinograms (ERGs) can be obtained. It is known that during light adaptation considerable changes occur in the cone ERGs of man, monkeys and mice. All these species have vascular retinae. In the present study we examined whether the same applies to mammalian species with a limited retinal vasculature (rabbits) or avascular retinae (guinea pigs), and which both have two types of cones but scotopic ERGs with completely different morphology. Material and methods: ERGs were recorded from anaesthetized rabbits and guinea pigs with corneal electrodes made from steal wire. Copper wire placed in the mouth of the animal served as reference electrode, and a subcutaneous needle as ground. Recordings were amplified 1000-fold, with bandwidth settings at 1-1000 Hz, and fed into a computer via an A/D converter. Corneas were anaesthetized with a topical application of proparacaine, and pupils dilated with topical application of tropicamide. ERGs were elicited with brief (10 msec) light flashes, and the retina light adapted with a steady white background light. Results: The scotopic b-wave is more than twice the amplitude of the a-wave in rabbits, while the scotopic b-wave in guinea pigs is only slightly larger than the a-wave. The b-wave of the cone ERG is twice the amplitude of the cone a-wave in both species. Once a background light has been turned on, the amplitude increases in both species and the process of light adaptation reaches a peak about 10 minutes thereafter. The b-wave implicit time is shortened by light adaptation in rabbits, but not in guinea pigs. Oscillatory potentials are present in guinea pig ERGs when recorded in dark but not when recorded in light. Conclusions: Mammals that have avascular retinae and which are without long-wavelength cones show evidence of light adaptation of the cone ERG. In guinea pigs the cone ERG increases in amplitude during light adaptation without concomitant shortening of the implicit time. These changes occur at similar rate in rabbits and guinea pigs. The oscillatory potentials in rabbits increase in amplitude but not in guinea pigs. These results suggest that different mechanisms determine the light adaptation of the cone ERG in guinea pigs than in rabbits. Key words: ERG, avascular retina, cones, light adaptation, rabbit, guinea pig. Correspondence: Þór Eysteinsson. E-mail: thoreys@hi.is |
Inngangur
Þegar sjónhimna er aðlöguð að bakgrunnsljósi, er hægt að skrá sjónhimnurit keilna (photopic electroretinogram, ERG) og nota sem mælikvarða á næmi augans fyrir birtu. Sjónhimnurit er rafsvar sjónhimnu sem vakið er með ljósblikkum, og líkt og hjartarafrit endurspeglar rafvirkni vefsins í heild. Sjónhimnurit er hægt að skrá með skráningarskauti staðsettu á hornhimnu og með því er hægt að fá raflífeðlisfræðilegt mat á ástandi stafa og keilna. Hreint sjónhimnurit keilna, án innleggs frá stöfum í svari, fæst ef stöðugt bakgrunnsljós er látið skína á augað sem skráð er frá. Spenna sjónhimnurits hækkar og dvöl þess styttist frá því kveikt er á bakgrunni og næstu 5-15 mínútur þar á eftir. Sýnt hefur verið fram á þetta meðal annars í mönnum (1,2) og öpum (3) sem hafa tvær til þrjár gerðir keilna og svipað litrófsnæmi. Einnig hefur þetta verið skoðað í músum (4) sem hafa tvær gerðir keilna, en sérstakt litrófsnæmi þar sem önnur gerðin er næm á útfjólubláa sviðinu (5). Þessar tegundir spendýra hafa æðar í sjónhimnu. Í sjónhimnuriti eru nokkrar afmarkaðar bylgjur, sem fram koma í svarinu á tilteknum tíma eftir ljósertingu. Fyrst kemur fram neikvæð sveifla í spennu sem kallast a-bylgja, og talin orsakast af rafvirkni ljósnema fyrst og fremst. Því næst kemur fram jákvæð bylgja sem kallast b-bylgja og talin er orsakast af rafvirkni taugafrumna og Muller-stoðfrumna (glia) í sjónhimnu. Við ljósaðlögun kemur aukning í spennu fyrst og fremst fram í b-bylgju og í svari við blikkandi ljósáreitum, en einnig stundum í a-bylgju, sem er þó háð birtumagni. Aukningin er mest þegar notað er mikið birtumagn (2) sem mettar stafi, en það eru þær aðstæður sem eru notaðar klínískt til að hámarka keilusvör. Þau ferli sem liggja að baki aukinni spennu sjónhimnurits í ljósaðlögun eru ekki að fullu þekkt, en tvö möguleg ferli koma til greina. Annars vegar er margt sem bendir til þess að það sem liggi til grundvallar þessum áhrifum einskorðist aðeins við keilukerfið og hlutverk Ca2+ í ljósnemum. Við ljósaðlögun orsakar minnkun innflæðis Ca2+ inn í keilur endurafskautun þeirra (6,7) og þessari endurafskautun fylgir aukning í svari þeirra. Þegar bakgrunnsljós er kveikt verður yfirskautun í himnuspennu keilna vegna vatnsrofs á cGMP sem heldur jónagöngum opnum í myrkri. Með meiri ljósaðlögun afskautast keilur aftur, stig af stigi, vegna almennrar stjórnunar Ca2+ á styrk cGMP í ytri liðum ljósnema í gegnum neikvæða endurgjöf. Hins vegar gæti verið um einhvers konar áhrif mögnunar að ræða við taugamót í ytra flókalagi (outer plexiform layer). Með skráningum á himnuspennu með innanfrumumælingum hefur verið sýnt fram á breytingar í mögnun sem verða á taugamótum keilna og láréttra frumna við ljósaðlögun (8). Svör láréttra frumna stækka en ekki svör keilna. Ekki er ljóst hvort þessar breytingar í mögnun eru orsök breytinga í sjónhimnuriti keilna við ljósaðlögun, en tímaferill er mjög svipaður.
Vegna þeirra breytinga sem verða í keilu sjónhimnurits spendýra var athugað hvort sama eigi við um tegundir spendýra sem hafa takmarkað magn af æðum í sjónu (kanínur) eða engar æðar (marsvín) (9), og annað litrófsnæmi. Báðar dýrategundir eru með tvær tegundir keilna í sjónhimnu, stuttbylgukeilur (bláar) og miðbylgjukeilur (grænar) (5). Langbylgjukeilur (rauðar) eru í flestum þeim tegundum sem sýna breytingar í sjónhimnuriti keilna við ljósaðlögun, þótt menn með skerta starfsemi langbylgjukeilna (protanopia) sýni þær einnig (10). Ekki er þó vitað nákvæmlega um framlag einstakra keilna í ljósaðlögun. Hér var athugað hvort sjónhimna án rauðra keilna og án æða sýni sams konar ljósaðlögun og í spendýrum sem hafa æðar í sjónhimnu og rauðar keilur. Jafnframt var markmið þessarar rannsóknar að bera saman ljósaðlögun keilna í sjónhimnu kanína og marsvína.
Efniviður og aðferðir
Sjónhimnurit var skráð frá fimm marsvínum og fimm kanínum. Dýrin voru aðlöguð að rökkri í 30 mínútur fyrir tilraun og undirbúin við dimmt rautt ljós sem dregur á engan hátt úr ljósnæmi. Ketamín var notað til svæfingar og því sprautað undir húð. Hornhimna var staðdeyfð með 0,5% próparakaíni og sjáaldur víkkað með 1% trópikamíði. Saltupplausn var dreypt í auga með reglulegu millibili til að koma í veg fyrir þornun hornhimnu. Skráningarskaut úr þunnum vír úr ryðfríu stáli, húðuðum með 1% methýlsellulósa til að auka leiðni, var staðsett á hornhimnu augans. Vírinn er vafinn í vafning með 3-5 mm þvermál, til að falla vel að hornhimnu (11). Koparvír var notaður sem viðmiðunarskaut og hann staðsettur í munni dýrsins. Sjónhimnuritssvör voru mæld milli þessara tveggja skauta. Platínunál var notuð sem jarðtenging og henni stungið undir húð á kviði. Sjónhimnuritssvör voru vakin með 10 µs hvítu (xenon) ljósblikki (1,8 log cd (candella) x sek/m2) frá Grass PS-33 ljósertara (Astro-Med/Grass Inc., USA), fyrst í rökkri, og síðan eftir að kveikt hafði verið á björtu bakgrunnsljósi (1,7 log cd/m2). Sjónhimnuritssvör við þessar aðstæður endurspegla svör keilna. Ýmist var notað eitt ljósblikk eða 20-30 riða blikkandi áreiti. Gögnum var safnað á um 30 sekúndna fresti í 15-20 mínútur. Úttakið frá skautunum var leitt inn í Digitimer NeuroLog (Digitimer Ltd., England) magnara (x 1000 mögnun) og rafsíur stilltar á bandvídd 1-1000 Hz. Frá mögnurum voru gögnin tekin inn á Macintosh tölvu með A/D breytikorti (MacADIOS Superscope II, GW Instruments, USA) og spennuvídd og dvöl sjónhimnurits mæld. Meðan á tilraunum stóð voru dýrin látin liggja á einangraðri plötu með yfirborðshita um 37 gráður.Unnið var úr gögnum með tölfræðiforritinu Instat (útgáfa 2.01). Beitt var pöruðu t-prófi við tölfræðilega greiningu og miðað við marktektarmörk a = 0,05 (öryggismörk 95%). Spennuútslag rafsvara er gefið upp sem meðaltal ± staðalvilla (SEM).
Niðurstöður
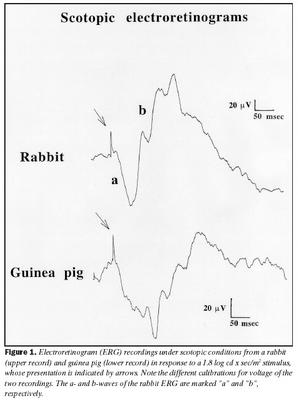
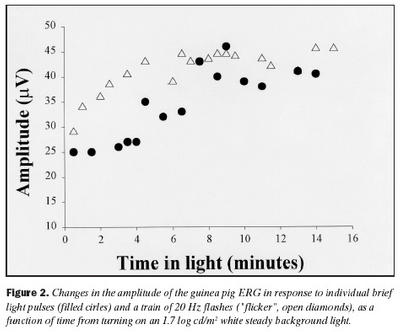
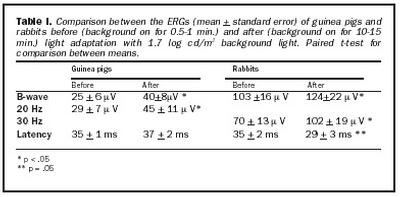
Áður en áhrif ljósaðlögunar voru mæld voru sjónhimnurit í rökkri (scotopic ERG) skráð. Mynd 1 sýnir að eftir aðlögun að rökkri eru sjónhimnurit kanínu og marsvíns ólík að lögun, en sýna bæði a- og b-bylgjur eins og sýnt er á myndinni. Sjónhimnurit kanínu er svipað og hjá mönnum, músum og öpum, með stóra a-bylgju, en enn stærri b-bylgju (b/a hlutfall = 2,3 að meðaltali). Aftur á móti er sjónhimnurit marsvína mjög lágt að spennu og næstum neikvætt, hefur stóra a-bylgju en aðeins eilítið stærri b-bylgju (b/a hlutfall =1,3 að meðaltali).Þegar áhrif ljósaðlögunar voru skoðuð kom í ljós að sjónhimnurit keilna hjá marsvínum eykst að spennu við aðlögun að ljósi, en er háð tíðni ertingar (mynd 2). Ef tíðni ertingar er hærri en 20 Hz lækkar spenna sjónhimnurits hjá marsvínum verulega og er nánast ómælanlegt við 30 Hz. B-bylgja sem svar við stuttum ljósblikkum var að meðaltali 25 ± 6 µV rétt eftir að kveikt hafði verið á bakgrunnsljósi, en eftir um 10 mínútna aðlögun að ljósinu jókst spennan að meðaltali í 40 ± 8 µV. Þessi munur í spennu er tölfræðilega marktækur (p = ,0178).
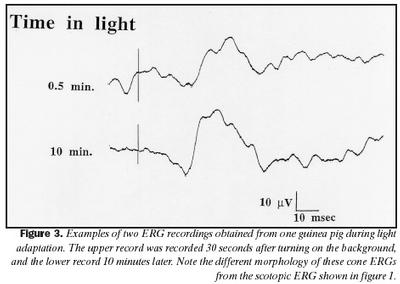
Svar við 20 Hz ljósblikkum var að meðaltali 29 ± 7 µV rétt eftir að kveikt hafði verið á bakgrunnsljósi, en jókst að spennu í 45 ± 11 µV að meðaltali eftir um 10 mínútna aðlögun að ljósi, og er þessi munur marktækur (p = ,0236). Aukning í spennu b-bylgju var svipuð og í spennu svars við 20 Hz ljósblikkum við aðlögun að ljósi. Það er hins vegar eftirtektarvert af mynd 2 að aukningin í svari við 20 Hz ljósblikkum fylgir veldisfalli, en ekki b-bylgja. Á mynd 3 eru sýndar tvær skráningar sjónhimnuritssvara frá marsvíni við stuttum ljósblikkum 30 sekúndum eftir að kveikt var á bakgrunnsljósi og eftir 10 mínútna stöðuga aðlögun að sama ljósi.
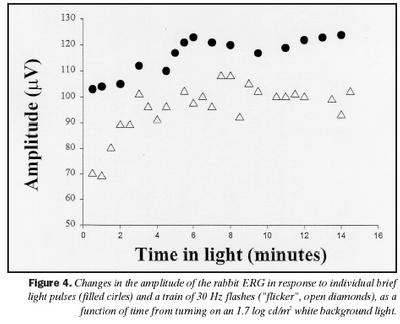
Sjónhimnurit keilna er stærri að spennu í kanínum en marsvínum. Sjónhimnurit keilna hjá kanínum eykst að spennu við aðlögun að ljósi, en er háð tíðni ertingar. Sjónhimnurit keilna hjá kanínum er mælanlegt með tíðni ertingar allt að 50 Hz. Mynd 4 sýnir breytingar í spennu sjónhimnurits sem fall af tíma frá því kveikt var á bakgrunnsljósi fyrir kanínur. B-bylgja sem svar við stuttum ljósblikkum var að meðaltali 103 ± 16 µV en jókst við ljósaðlögun í 124 ± 22 µV að meðaltali. Þessi munur í spennu er tölfræðilega marktækur (p = ,0221).
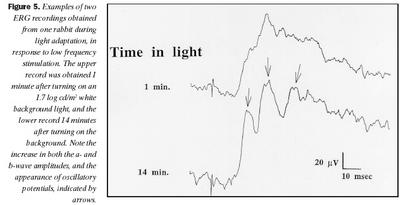
Svar við 30 Hz ljósblikkum var að meðaltali 70 ± 13 µV en jókst við aðlögun að ljósi í 102 ± 19 µV að meðaltali, sem er marktækur munur (p = ,0273). Það er eftirtektarvert, að aukning í svari við 30 Hz ljósblikkum (32 mV) var meiri en í b-bylgju (21 mV) við aðlögun að ljósi, og fylgdi veldisfalli, en ekki b-bylgja. Mynd 5 sýnir tvær skráningar frá kanínu við stuttum ljósblikkum, einni mínútu eftir að kveikt var á bakgrunni og eftir 14 mínútna stöðuga aðlögun að bakgrunnsljósi. Þrjár aðgreinanlegar sveifluspennur birtust í svörum um tveimur mínútum eftir að kveikt var á bakgrunnsljósi og jukust að spennu með aukinni aðlögun að birtu. Sambærilegar breytingar í sveifluspennum voru ekki greinanlegar í sjónhimnuriti keilna hjá marsvínum, eins og sjá má af mynd 3.
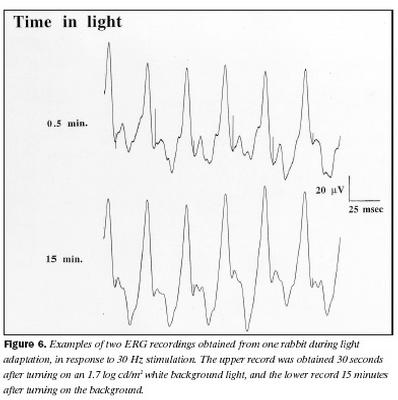
Dæmi um svör kanína við 30 Hz ljósblikkum, hálfri mínútu eftir að kveikt var á bakgrunni og eftir 15 mínútna stöðuga aðlögun að bakgrunnsljósi eru sýnd á mynd 6. Svör við 30 Hz ljósertingu sýndu tvo aðgreinanlega þætti, sem jukust báðir að spennu við aðlögun að ljósi, eins og sjá má af mynd 6.
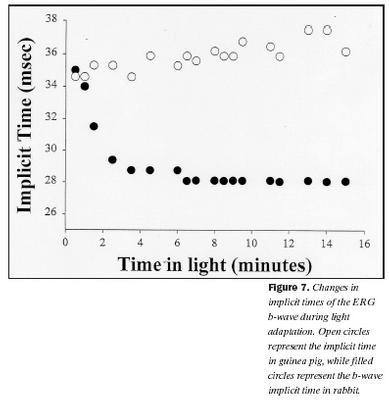
Dvöl (implicit time) b-bylgju styttist ekki hjá marsvínum og hélst að mestu óbreytt við ljósaðlögun og var að meðaltali um 36 ± 2 msek. Aftur á móti styttist dvölin hjá kanínum svipað og sést hjá öðrum spendýrum og var að meðaltali 35 ± 2 msek í upphafi, en við aðlögun að ljósi styttist hún að meðaltali í 29 ± 3 msek. Þessi munur er tölfræðilega marktækur (p = ,05). Mynd 7 sýnir dvöl sem fall af tímanum frá því kveikt var á bakgrunnsljósi fyrir bæði kanínur og marsvín.
Í töflu I eru dregnar saman niðurstöður, til samanburðar á sjónhimnuritssvörum frá kanínum og marsvínum.
Umræða
Niðurstöður okkar benda til að sjónhimnurit keilna í marsvínum og kanínum aukist að spennu við aðlögun sjónhimnu að ljósi, í samræmi við fyrri rannsóknir (1-4) á spendýrum sem hafa æðar í sjónhimnu, en annað litrófsnæmi keilna. Þessar breytingar í sjónhimnuriti við ljósaðlögun virðast því verða óháð því hvort æðakerfi er í sjónhimnu eða ekki, og því hvert er litrófsnæmi keilna í viðkomandi spendýri. Sjónhimnurit keilna er stærra að spennu í kanínum en marsvínum. Hins vegar eykst b-bylgja í sjónhimnuriti keilna hlutfallslega meira í marsvínum við ljósaðlögun.Dvöl b-bylgju í keilusvari marsvína styttist ekki við aðlögun að ljósi. Aftur á móti styttist dvöl keilusvara hjá kanínum við aðlögun að ljósi, eins og í mönnum, músum og öpum, og er það í samræmi við hraðari svörun keilukerfis heldur en stafakerfis. Dvöl styttist mest fyrstu mínútu eftir að kveikt hefur verið á bakgrunnsljósi og áframhaldandi aðlögun hefur takmörkuð áhrif fremur í þá veru að stytta dvöl. Hjá mönnum er misjafnt hvernig og hvort dvölin styttist eftir því hversu björt áreiti eru notuð (12). Það kann að skýra hvers vegna dvölin styttist ekki hjá marsvínum, þar sem þær eru kannski næmari fyrir önnur ljósáreiti, en í þessari rannsókn var aðeins notaður einn ljósstyrkur áreitis til að vekja sjónhimnurit keilna.
Svar við stuttum ljósblikkum vekur afmarkaðar, aðgreinanlegar bylgjur í sjónhimnuriti og við aðlögun að ljósi eru það keilur sem miðla ljóssvöruninni. Við háa tíðni ertingar, 20-30 rið í þessari rannsókn, eru það einnig keilur sem miðla svöruninni en ekki er hægt að aðgreina a- og b-bylgjur í því svari. Í báðum tilvikum var notað sams konar stöðugt bakgrunnsljós til að útiloka stafi og aðlaga keilur að ljósi. Samt sést nokkur munur í aðlögunarferli b-bylgju og sjónhimnuritssvars við hárri tíðni ertingar, fyrst og fremst sá að við háa tíðni ertingar fylgir aðlögunarferlið veldisfalli hjá báðum tegundum, en ekki b-bylgja. Einfaldasta skýringin á þessum mun er sú, að það sjónhimnurit sem er vakið með stuttum ljósblikkum á sér að einhverju leyti aðrar uppsprettur en það sjónhimnurit sem vakið er með hárri tíðni ljósertingar. Talið er víst að b-bylgja, vakin með stuttum ljósblikkum, eigi sér að hluta uppsprettu í himnuspennubreytingum Muller-frumna, en þar sem þær himnuspennubreytingar eru mjög hægar er mjög ólíklegt að þær séu þáttur í sjónhimnuritssvari við 20-30 riða ljósertingu (13).
Ljósaðlögun hefur mun meiri áhrif á sveifluspennur hjá kanínum en marsvínum. Í upphafi aðlögunar að ljósi eru sveifluspennur hjá hvorugri tegund, þrjár slíkar birtast síðar hjá kanínum og stækka að spennu og ná hámarksspennu eftir um 15 mínútur í birtu. Þessar sveifluspennur eru vegna ljóssvörunar keilna, en talið er að rafvirkni flókafrumna (amacrine cells) og milliflókafrumna (interplexiform cells) séu raflífeðlisfræðileg uppspretta þeirra (14,15). Einfaldasta skýringin á þeim muni sem er á áhrifum ljósaðlögunar á sveifluspennur í þessum tegundum er að starfsemi þessara frumna sé ólík í kanínum og marsvínum, eða taugatengsl við keilur séu ólík. Niðurstöður þessarar rannsóknar benda því til að bæði kanínur og marsvín séu raunhæf líkön til grunnrannsókna á ljósaðlögun, og geti hvort með sínum hætti gefið nánari upplýsingar um þau ferli sem liggja að baki.
Þau líffræðilegu ferli sem eiga sér stað í sjónhimnunni við ljósaðlögun eru ekki að fullu þekkt, en munur á áhrifum ljósaðlögunar á spennu og dvöl sjónhimnurits keilna í marsvínum og kanínum bendir til þess að ólíkir ferlar liggi þar að baki. Það gæti orsakast af einhverjum mun í lífeðlislegum ferlum í keilum þessara tegunda eða mun í ferlum í taugamótum af því tagi sem fundist hafa (8) með innanfrumuskráningum í froskdýrum. Ein leið til að prófa þessar tilgátur er að breyta starfsemi taugamóta í sjónhimnu lyfjafræðilega, og athuga hvort ljósaðlögun sjónhimnurits keilna breytist við það. Eru slíkar athuganir nú í gangi á rannsóknarstofu okkar.
Klínísk gagnsemi sjónhimnuritsmælinga við ljósaðlögun felst í mögulegu notagildi þeirra í greiningu á ákveðnum augnsjúkdómum. Samkvæmt alþjóðlegum staðli fyrir skráningu sjónhimnurits frá fólki, ber að aðlaga að ljósi í minnst 10 mínútur til að fá raunhæfa raflífeðlisfræðilega mynd af starfsemi keilna (16). Samt geta breytingarnar í sjónhimnuriti við ljósaðlögun sem slíkar gefið vísbendingar um starfsemi keilna í sjúkdómum. Ljóst er að breytingar í starfsemi stafa er ekki orsakaþáttur (12). Vitað er að í sjónufreknum (retinitis pigmentosa) er ljósaðlögun sjónhimnurits keilna, ef það svar er fyrir hendi, óeðlilega hæg og tekur lengri tíma að ná hámarki (6,17). Í kynbundnu sjónurofi (retinoschisis) er ljósaðlögun sjónhimnuritssvars við 30 riða áreitum eðlileg (18), en engin ljósaðlögun sjónhimnurits keilna mælist ef notuð er lægri ertingartíðni en 8 Hz (óbirt gögn höfunda). Er það hugsanlega vegna þess að Muller-frumur, sem eru þáttur í myndun b-bylgju við lága ertingartíðni, eru sundraðar í sjónurofi (19). Niðurstöður frá sjúklingum með sjúkdóma í miðgróf eru misvísandi; ljósaðlögun sjónhimnurits er eðlileg í Sjúkdómi Bests (18) en í sjúkdómi Stargardts (fundus flavimaculatus) ýmist óeðlileg eða ekki fyrir hendi (óbirt gögn höfunda). Ófullkomin (incomplete-type) meðfædd staðbundin náttblinda, (I-CSNB) er arfgengur augnsjúkdómur, þar sem sjónhimnurit stafa er "neikvætt", sem þýðir að a-bylgja er stærri að spennu en b-bylgja, en sjónhimnurit keilna er oft eðlilegt (18). Talið er að um sé að ræða óeðlileg taugatengsl milli stafa og keilna annars vegar og tvískautafrumna (bipolar cells) hins vegar. Sjúkdómurinn einkennist af náttblindu og rýru sjónhimnuriti í rökkri. Við ljósaðlögun með bakgrunni á sama hátt og notað var í þessari rannsókn, eftir rökkuraðlögun á undan, sýna þessir sjúklingar aftur á móti of-eðlilega (supernormal) aukningu í spennu við 30 riða blikkandi áreiti, þar sem spennan er mjög lítil í upphafi en eykst mikið við aðlögun að ljósi (18). Þessi of-eðlilega aukning í svörun og kerfisbundnu breytingar í lögun bylgja við aðlögun koma ekki fram í eðlilegri sjónhimnu og hafa ekki sést í öðrum sjúkdómum í sjónhimnu sem hafa verið skoðaðir. Hvort ástæðan fyrir of-eðlilegri ljósaðlögun sjónhimnurits í ófullkominni meðfæddri staðbundinni náttblindu felst í starfsemi keilna eða annarra taugafrumna í sjónhimnu er óljóst. Af öllu er þó ljóst að í ofangreindum sjúkdómum er munur í aðlögun keilna að ljósi. Þar sem sjónskerpa ræðst af starfsemi keilna er ástand þeirra mikilvægt atriði þegar meta á horfur varðandi sjón, til dæmis í hrörnunarsjúkdómum í sjónhimnu. Ljósaðlögun keilna tryggir að sjónskerpa haldist jöfn við mismunandi birtumagn í umhverfi. Niðurstöður okkar benda til að önnur ferli ráði ljósaðlögun sjónhimnurits keilna hjá marsvínum en kanínum.
Heimildir
1. Armington JC, Biersdorf WR. Long-term light adaptation of the human electroretinogram. J Comp Physiol Psychol 1958; 51:1-5.2. Peachey NS, Alexander KR, Fishman GA, Derlacki DJ. Properties of the human cone system electroretinogram during light adaptation. Appl Optics 1989; 28: 1145-50.
3. Murayama K, Sieving PA. Different rates of growth of monkey and human photopic a-, b-, and d-waves suggest two sites of ERG light adaptation. Clin Vis Sci 1992; 7: 385-92.
4. Peachey NS, Goto Y, Ubaidi MRA, Naash MI. Properties of the mouse cone-mediated electroretinogram during light adaptation. Neurosci Letters 1993; 162: 9-11.
5. Jakobs GH. The distribution and nature of color vision among the mammals. Biol Review 1993; 68: 413-71.
6. Gouras P, MacKay CJ. Light adaptation of the electroretinogram: diminished in retinitis pigmentosa. Invest Opthalm Vis Sci 1989; 30: 619-24.
7. Matthews HR, Fain GL, Cornwall MC. Role of cytoplasmic calcium concentration in the bleaching adaptation of salamander cone photoreceptors. J Physiol 1996; 490: 293-303.
8. Yang XL, Wu SM. Response sensitivity and voltage gain of the rod- and cone horizontal cell synapses in dark- and light adapted tiger salamander retina. J Neurophysiol 1996; 76: 3863-74.
9. Schaepdrijver LD, Simoens P, Lauwers H, Geest JPD. Retinal vascular patterns in domestic animals. Res Vet Sci 1989; 47: 34-42.
10. Eysteinsson Þ, Frumkes TE. Light adaptation of the cone system electroretinogram in dichromats and anomalous trichromats [abstract]. Invest Ophthalm Vis Sci 1995; 36: abstract 2067.
11. Goto Y. An electrode to record the mouse corneal electroretinogram. Doc Ophthalmol 1993; 91: 147-54.
12. Peachey NS, Alexander KR, Derlacki DJ, Fishman GA. Light adaptation, rods, and the human cone flicker ERG. Visual Neurosci 1992; 8: 145-50.
13. Miller RF, Dowling JE. Intracellular responses of the Muller cells of mudpuppy retina: their relation to the b-wave of the ERG. J Neurophysiol 1970; 33: 323-41.
14. Arnarsson Á, Einarsson JM, Eysteinsson Þ. Sveifluspennur í sjónhimnu: Áhrif GABA-agonista. Læknablaðið 1995; 81: 412-6.
15. Wachtmeister L. Oscillatory potentials in the retina: what do they reveal. Progr Ret Eye Res 1998; 17: 485-521.
16. Marmor MF, Zrenner E. Standard for clinical electroretinography (1994 update). Doc Ophthalmol 1995; 89: 199-210.
17. Miller S, Sandberg MA. Cone electroretinographic change during light adaptation in retinitis pigmentosa. Invest Ophthalm Vis Sci 1991; 32: 2536-41.
18. Miyake Y, Horiguchi M, Ota I, Shiroyama N. Characteristic ERG flicker anomaly in incomplete CSNB. Invest Ophthalm Vis Sci 1987; 28: 1816-23.
19. Condon GP, Brownstein S, Wang NS, Kearns AF, Ewing CC. Congenital hereditary (juvenile x-linked) retinoschisis. Histopathological and ultrastructural findings in three eyes. Arch Ophthalmol 1986; 104: 576-83.